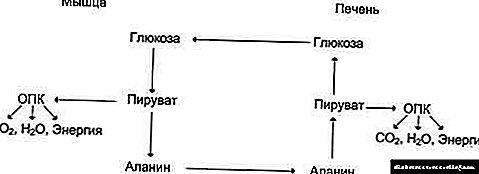
Xolesterin biosintezi va uning biokimyosi - diabet
Shubhasiz, xolesterin keng jamoatchilikka ma'lum bo'lgan lipid bo'lib, u qonda yuqori xolesterin va inson yurak-qon tomir kasalliklarining chastotasi o'rtasidagi bog'liqlik tufayli mashhurdir. Xolesterolni hujayra membranalarining tarkibiy qismi va steroid gormonlari va safro kislotalari uchun muhim omil sifatida hal qiluvchi rolga kamroq e'tibor qaratildi. Xolesterol ko'plab hayvonlarga, shu jumladan odamlarga kerak, ammo uning sutemizuvchi oziq-ovqatda bo'lishi majburiy emas - tana hujayralari o'zlari buni oddiy prekursorlardan sintez qilishlari mumkin.
Ushbu 27-uglerodli birikmaning tuzilishi uning biosintezi uchun murakkab yo'lni taklif qiladi, ammo uning barcha uglerod atomlari bitta prekursor - atsetat tomonidan ta'minlanadi. Izopren bloklari - Atsetatdan xolesterolgacha bo'lgan eng muhim vositachilar, ular ko'plab tabiiy lipidlarning prekursorlari bo'lib, izopren bloklari polimerizatsiya qilingan mexanizmlar barcha metabolik yo'llarda o'xshashdir.
Atsetatdan xolesterin biosintezi yo'lidagi asosiy bosqichlarni o'rganishdan boshlaymiz, so'ngra xolesterolni qon oqimi orqali tashish, uning hujayralar tomonidan so'rilishi, xolesterin sintezining normal holati va buzilgan assimilyatsiya yoki tashishni tartibga solish masalalarini muhokama qilamiz. Keyin biz safro kislotalari va steroid gormonlari kabi xolesteroldan kelib chiqadigan boshqa moddalarga qaraymiz. Va nihoyat, ko'plab birikmalar hosil bo'lishining biosintetik yo'llarining tavsifi - xolesterol sintezi bilan umumiy erta bosqichlar bo'lgan izopren bloklari hosilalari biosintezda izoprenoid kondensatsiyasining g'ayrioddiy xususiyatlarini namoyish etadi.
Xolesterol atsetil-CoAdan to'rt bosqichda ishlab chiqariladi
Xolesterol, uzoq zanjirli yog 'kislotalari singari, asetil-CoA-dan ishlab chiqariladi, ammo yig'ilish shakli butunlay boshqacha. Birinchi tajribalarda 14 S ga teng bo'lgan atsetat metil yoki karboksil uglerod atomi hayvonlar ozuqasiga qo'shildi. Yorliqning ikki guruh hayvonlardan ajratilgan xolesterolda tarqalishiga asoslanib (21-32-rasm), xolesterin biosintezi fermentativ bosqichlari tavsiflangan.
Shakl 21-32. Xolesterol uglerod atomlarining manbai. Metil uglerod (qora) yoki karboksil uglerod (qizil) bilan belgilangan radioaktiv atsetat yordamida tajriba davomida aniqlangan. Kondensatsiyalangan strukturada halqalar A dan D harflari bilan belgilanadi.

Rasmda ko'rsatilgandek sintez to'rt bosqichda amalga oshiriladi. 21-33: (1) uchta asetat qoldiqlarining kondensatsiyasi mevalonatning oltita uglerodli oralig'ini hosil qiladi, (2) mevalonatning faollashtirilgan izopren bloklariga aylantirilishi, (3) 30 karbonli chiziqli skalenni hosil qilish uchun oltita besh uglerodli izopren birliklarining polimerizatsiyasi, (4) skalolning tsikllanishi. steroid yadrosining to'rt halqasi, undan keyin xolesterin hosil bo'lishi bilan bir qator o'zgarishlar (oksidlanish, metil guruhlarining ko'chishi yoki ko'chishi).
Shakl 21-33. Xolesterin biosintezi haqida umumiy rasm. Matnda sintezning to'rtta bosqichi muhokama qilinadi. Skalendagi izopren bloklari qizil chiziqli chiziqlar bilan belgilanadi.

Bosqich (1). Asetatdan mevalonat sintezi. Xolesterol biosintezining birinchi bosqichi oraliq mahsulot hosil bo'lishiga olib keladi mevalonate (21-34-rasm). Ikki asetil CoA molekulasi oltita uglerodli birikma hosil qilish uchun uchinchi asetil CoA molekulasi bilan kondensatsiyalanadigan asetoatsetil CoA berish uchun kondensatsiyalanadi β-gidroksi-β-metglutaryil-CoA (HM G -CoA). Ushbu ikkita birinchi reaktsiya katalizlanadi tiolaza navbati bilan NM G -CoA sintazasi. Sitozolik NM G-CoA sintazasi Ushbu metabolik yo'l keton jismlarni shakllantirish jarayonida NM G -CoA sintezini katalizlovchi mitoxondrial izoenzimdan farq qiladi (17-18-rasmga qarang).
Shakl 21-34. Atsetil-CoA dan mevalonatning shakllanishi. Atsetil-CoA dan olingan C-1 va C-2 mevalonat manbai pushti rang bilan ta'kidlangan.

Uchinchi reaktsiya butun jarayonning tezligini cheklaydi. Unda NM G -CoA mevalonatga kamayadi, buning uchun har ikki NA D PH molekulasi ikkita elektronni ta'minlaydi. HMG-CoA reduktazasi - silliq ER ning ajralmas membrana oqsili bo'lib, u keyinchalik ko'rib chiqamiz, xolesterin hosil bo'lishining metabolik yo'lini tartibga solishning asosiy nuqtasi bo'lib xizmat qiladi.
Bosqich (2). Mevalonatning ikkita faollashtirilgan izoprenga aylanishi. Xolesterol sintezining keyingi bosqichida ATF molekulalaridan mevalonatga uchta fosfat guruhi yuboriladi (21-35-rasm). 3-fosfo-5-pirofosfomevalonat oralig'idagi C-3 mevalonatidagi gidroksil guruhiga bog'langan fosfat yaxshi tarkibi hisoblanadi, keyingi bosqichda ushbu fosfatlar ikkalasi ham, qo'shni karboksil guruhining tarkibi besh karbonli mahsulotda ikki tomonlama aloqani hosil qiladi - 3 -izopentenil pirofosfat. Bu faollashtirilgan ikkita izoprenning birinchisi - xolesterin sintezining asosiy ishtirokchilari. Δ 3 -isopentenilpirofosfatning izomerlanishi ikkinchi faollashtirilgan izoprenni beradi dimetililil pirofosfat. O'simlik hujayralari sitoplazmasida izopentenil pirofosfat sintezi bu erda tasvirlangan yo'l bo'yicha sodir bo'ladi. Ammo o'simlik xloroplastlari va ko'plab bakteriyalar mevalonatdan mustaqil bo'lgan yo'ldan foydalanadilar. Ushbu alternativ yo'nalish hayvonlarda topilmaydi, shuning uchun yangi antibiotiklar yaratishda jozibali bo'ladi.
Shakl 21-35. Mevalonatni faollashtirilgan izopren bloklariga aylantirish. Faollashtirilgan oltita birlik birlashib, skalene hosil qiladi (21-36-rasmga qarang). 3-fosfo-5-pirofosfomevalonatning tarkibi pushti rangda ta'kidlangan. Kvadrat qavs ichida faraz oraliq bo'ladi.

Bosqich (3). Svkaleni hosil qilish uchun oltita faollashtirilgan izopren birliklarining kondensatsiyasi. Endi izopentenil pirofosfat va dimetilillil pirofosfat bosh pog'onali kondensatsiyaga uchraydi, bunda bitta pirofosfat guruhi harakatlanadi va 10 uglerodli zanjir hosil bo'ladi - geranil pirofosfat (21-36-rasm). (Pyrofosfat boshga yopishadi.) Geranil pirofosfat izopentenil pirofosfat bilan 15 ta uglerod oraliq shaklidagi quyma kondensatsiyaga uchraydi. farnesil pirofosfat. Va nihoyat, farnesil pirofosfatning ikki molekulasi "boshdan boshga" birlashadi, ikkala fosfat guruhi ham olib tashlanadi - hosil bo'ladi skalen.
Shakl 21-36. Skalen shakllanishi. 30 uglerod atomini o'z ichiga olgan skalen tuzilishi izopren (besh karbonli) bloklar tomonidan faollashtirilgan kondensatsiya paytida yuzaga keladi.

Ushbu vositachilar uchun umumiy nomlar ular avval izolyatsiya qilingan manbalarning nomlaridan kelib chiqadi. Atirgul moyining tarkibiy qismi bo'lgan Geraniol, geranium lazzatiga ega va akasiya farnesasining ranglarida topilgan farnesol vodiyning xushbo'y hidli nilufarga ega. Ko'pgina tabiiy o'simlik hidlari izopren bloklaridan yasalgan birikmalarga tegishli. Avval akula jigaridan ajratib olingan skvallen (Squalus turlari) 30 uglerod atomidan iborat: asosiy zanjirda 24 atom va metall o'rnini bosadigan oltita atom.
Bosqich (4). Skalenni steroid yadrosining to'rt halqasiga aylantirish. Shaklda 21-37 aniq ko'rinib turibdiki, skalen zanjirining tuzilishi va sterollar - tsiklik. Barcha sterollarda steroid yadrosini tashkil etadigan to'rtta kondensatsiyalangan halqalar mavjud va ularning barchasi C-3 atomida gidroksil guruhi bo'lgan spirtdir, shuning uchun inglizcha sterol deb ataladi. Harakat ostida skalen monooksigenasi O dan bitta kislorod atomi skalol zanjirining oxiriga qo'shiladi 2 va epoksid hosil bo'ladi. Ushbu ferment yana bir aralash funktsional oksidaza (qo'shimcha 21-1), NADPH yana kislorod atomini O-ga qaytaradi 2 ga H2 O. Mahsulotni ikki marta bog'lash skalen-2,3-epoksid Ajoyib izchil reaktsiya skalen epoksidi zanjirini tsiklik strukturaga aylantirishi uchun shunday joylashtirilgan. Hayvonlar hujayralarida bu siklizatsiya shakllanishiga olib keladi lanosterol Ukol yadrosiga xos bo'lgan to'rt halqani o'z ichiga oladi. Natijada, lanosterol taxminan 20 ta reaktsiyalar orqali xolesterolga aylanadi, bu ba'zi metal guruhlarining migratsiyasini va boshqalarini olib tashlashni o'z ichiga oladi. Taniqlilar orasida eng qiyinlaridan biri bo'lgan bu ajoyib biosintez yo'lining tavsifi Konrad Bloch, Teodor Linen, Jon Kornfort va Jorj Popiak tomonidan 1950 yillarning oxirlarida yaratilgan.
Shakl 21-37. Ringni yopish chiziqli skalenni kondensatsiyalangan steroid yadrosiga aylantiradi. Birinchi bosqich aralash koksubstrati N AD PH bo'lgan aralash funktsiyasi (monooksigenaza) bilan oksidaza katalizlanadi. Mahsulot epoksid bo'lib, keyingi bosqichda u steroid yadrosini hosil qiladi. Hayvonlar hujayralarida bu reaktsiyalarning yakuniy mahsuloti xolesterin bo'lib, boshqa organizmlarda undan biroz farq qiladigan sterollar hosil bo'ladi.


Xolesterol hayvonlar hujayralari, o'simliklar, zamburug'lar va protistlar uchun sterol xususiyatga ega bo'lib, boshqa shunga o'xshash sterollarni ishlab chiqaradi.
Ular skalen-2,3-epoksid uchun bir xil sintez yo'lidan foydalanadilar, ammo keyin yo'llar biroz ajralib ketadi va boshqa sterollar hosil bo'ladi, masalan, ko'plab o'simliklarda sigmasterol va zamburug'lardagi ergosterol (21-37-rasm).
21-1-misol. Skvalen sintezi uchun energiya xarajatlari
Bitta skalen molekulasini sintez qilish uchun energiya qiymati (ATP molekulalari sifatida ifodalangan) qancha?
Qaror. Atsetil-CoA dan skalen sintezida ATP faqat mevalonat faollashtirilgan izopren skalene prekursoriga aylantirilgan bosqichda sarflanadi. Skalol molekulasini yaratish uchun oltita faollashtirilgan izopren molekulasi, har bir faollashgan molekulani hosil qilish uchun uchta ATP molekulasi kerak. Hammasi bo'lib, bitta skalen molekulasini sintez qilish uchun 18 ATP molekulasi sarflanadi.
Tanadagi xolesterol aralashmalari
Umurtqali hayvonlarda jigarda ko'p miqdordagi xolesterin sintezlanadi. U erda sintezlangan xolesterolning bir qismi gepatotsitlarning membranalarida to'plangan, ammo u asosan uning uchta shaklidan biriga: safro (safro) xolesterol, safro kislotalari yoki xolesterin esterlariga eksport qilinadi. Safro kislotalari ularning tuzlari jigarda sintezlanadigan va lipidlarning hazm bo'lishiga hissa qo'shadigan xolesterolning hidrofilik hosilasidir (17-1-rasmga qarang). Xolesterolni aniqlovchilar jigarda harakat orqali hosil bo'ladi akil-CoA-xolesterol-asiltransferaza (ACAT). Ushbu ferment yog 'kislotasi qoldig'ini A koenzimidan xolesterin gidroksil guruhiga o'tkazishni kataliz qiladi (21-38-rasm), xolesterolni yanada hidrofobik shaklga aylantiradi. Sekretsiya qilingan lipoprotein zarralaridagi xolesterin esterlari xolesteroldan foydalangan holda yoki boshqa jigarda saqlanadigan boshqa to'qimalarga etkaziladi.
Shakl 21-38. Xolesterin esterlarini sintez qilish. Eterifikatsiya xolesterolni saqlash va tashish uchun yanada ko'proq hidrofobik shaklga aylantiradi.

Xolesterol o'sayotgan hayvon organizmining barcha to'qimalariga membranalarni sintez qilish uchun zarurdir va ba'zi organlar (masalan, buyrak usti bezlari va jinsiy bezlar) xolesterolni steroid gormonlariga qarshi vosita sifatida ishlatadilar (bu haqda quyida muhokama qilinadi). Xolesterin, shuningdek, D vitamini uchun muhim omil hisoblanadi (10-20-rasmlarga, 1-rasmga qarang).
Xolesterin va boshqa lipidlar plazma lipoproteinlarini olib yuradilar
Triatsilgliserol va fosfolipidlar kabi xolesterin va xolesterin esterlari deyarli suvda erimaydi, ammo ular sintez qilingan to'qimadan ular saqlanadigan yoki iste'mol qilinadigan to'qimalarga o'tishi kerak. Ular qon shaklida orqali o'tkaziladi qon plazmasidagi lipoproteinlar - o'ziga xos tashuvchi oqsillarning makromolekulyar komplekslari (apolipoproteinlar) turli xil birikmalarda ushbu komplekslarda mavjud bo'lgan fosfolipidlar, xolesterin, xolesterin esterlari va triatsilgliserollar bilan.
Apolipoproteinlar ("apo" lipidsiz oqsilni anglatadi) lipidlar bilan birlashib, lipoprotein zarralarining bir nechta fraktsiyalarini hosil qiladi - markazda gidrofob lipidlari bo'lgan sferik komplekslar va sirtdagi hidrofilik aminokislotalar zanjirlari (21-39-rasm, a). Lipitlar va oqsillarning turli xil kombinatsiyalari bilan turli xil zichlikdagi zarralar hosil bo'ladi - chilomicronlardan yuqori zichlikdagi lipoproteinlarga qadar. Ushbu zarralarni ultratsentrifugatsiya bilan ajratish mumkin (21-1-jadval) va elektron mikroskopiya yordamida ko'z bilan ko'rish mumkin (21-39-rasm, b). Lipoproteinlarning har bir fraktsiyasi ma'lum bir funktsiyani bajaradi, bu sintez joyi, lipid tarkibi va apolipoprotein miqdori bilan belgilanadi. Inson qon plazmasida kamida 10 xil apolipoproteinlar topilgan (21-2-jadval), ularning o'lchamlari, o'ziga xos antikorlar bilan reaktsiyalari va lipoproteinlarning har xil sinflarida xarakterli tarqalishi. Ushbu protein tarkibiy qismlari lipoproteinlarni muayyan to'qimalarga yo'naltiruvchi yoki lipoproteinlar ustida ishlaydigan fermentlarni faollashtiradigan signal sifatida ishlaydi.
21-1-jadval. Insonning plazma lipoproteinlari
Tarkibi (massa ulushi,%)
r = 513,000). LDL zarrasida taxminan 1500 molekula xolesterin esterlari mavjud bo'lib, ularning atrofida 500 molekula xolesterin qobig'i, 800 molekula fosfolipidlar va bitta apoB-100 molekulasi bor. b - elektron mikroskop bilan ko'rinadigan lipoproteinlarning to'rtta sinfi (manfiy bo'lganidan keyin). Soat yo'nalishi bo'yicha, chap chap rasmdan boshlab: chylomicrons - diametri 50 dan 200 nm, PL O NP - 28 dan 70 nm gacha, HDL - 8 dan 11 nm gacha va LDL - 20 dan 55 nm gacha. Lipoproteinlarning xususiyatlari jadvalda keltirilgan. 21-2.

Chylomicrons, sek da eslatib o'tilgan. 17, oziq-ovqat triatsilgliserollarini ichakdan boshqa to'qimalarga o'tkazing. Bular eng katta lipoproteinlardir, ular eng past zichlikka va triatsilgliserollarning eng yuqori nisbiy tarkibiga ega (17-2-rasmga qarang). Chylomicrons ingichka ichakni qoplaydigan epitelial hujayralar ER-da sintezlanadi, so'ngra limfa tizimidan o'tib, chap subklaviya venasi orqali qon oqimiga kiradi. Xilomikron apolipoproteinlari tarkibida apoB-48 (bu turdagi lipoproteinlar uchun noyob), apoE va apoC-II mavjud (21-2-jadval). AroC-II yog 'to'qimalarining, yurak, skelet mushaklari va emizikli sut bezlarining kapillyarlarida lipoprotein lipazasini faollashtiradi, bu to'qimalarga erkin yog' kislotalarining kirib borishini ta'minlaydi. Shunday qilib, ksilomikronlar oziq-ovqat yog'li kislotalarini to'qimalarga o'tkazadi, u erda ular iste'mol qilinadi yoki yoqilg'i sifatida saqlanadi (21-40-rasm). Xylomikron qoldiqlari (asosan triatsilgliserollardan ajralib chiqadi, ammo baribir xolesterol, apoE va apoB-48) qon aylanishi orqali jigarga etkaziladi. Jigarda retseptorlari chilomikron qoldiqlari tarkibidagi apoE bilan bog'lanadi va ularning endotsitoz orqali so'rilishini ta'minlaydi. Gepatotsitlarda bu qoldiqlar tarkibidagi xolesterolni chiqaradi va lizosomalarda yo'q qilinadi.
21-2-jadval. Odam plazmasidagi lipoprotein apolipoproteinlar
Funktsiya (agar ma'lum bo'lsa)
L CAT-ni faollashtiradi, ABC tashuvchisi bilan o'zaro ishlaydi
L CAT ni inhibe qiladi
L CAT, xolesterolni tashish / tozalashni faollashtiradi
LDL retseptorlari bilan bog'lanadi
Chylomicrons, VLDL, HDL
Chylomicrons, VLDL, HDL
Chylomicrons, VLDL, HDL
VLDL va chilomikron qoldiqlarini tozalashni boshlaydi
Ovqat tarkibida hozirgi vaqtda yoqilg'i sifatida ishlatilishi mumkin bo'lgan miqdordan ko'proq yog 'kislotalari mavjud bo'lsa, ular jigarda triatsilgliserollarga aylanadi, ular o'ziga xos apolipoproteinlar bilan fraktsiyani hosil qiladi. juda past zichlikdagi lipoproteinlar (VLDL). Jigarda ortiqcha uglevodlarni triatsilgliserollarga aylantirish va VLDL sifatida eksport qilish ham mumkin (21-40-rasm, a).Triatsilgliserollarga qo'shimcha ravishda VLDL fraktsiyasida ma'lum miqdordagi xolesterin va xolesterin esterlari, shuningdek apoB-100, apoC-1, apoC-II, apoC III va apoE mavjud (21-2-jadval). Ushbu lipoproteinlar qon orqali jigardan mushak va yog 'to'qimalariga etkaziladi, bu erda lipoprotein lipaza apo-C II tomonidan faollashtirilgandan so'ng VLDL fraksiyasining triatsilgliserollaridan erkin yog' kislotalari ajralib chiqadi. Adipotsitlar bo'sh yog 'kislotalarini ushlaydi, yana triatsilgliserollarga aylantiradi, ular bu hujayralarda lipid qo'shimchalari (tomchilar), miotsitlar shaklida saqlanadi, aksincha energiya hosil qilish uchun darhol yog'li kislotalarni oksidlaydi. Ko'pincha VLDL qoldiqlari gepatotsitlar tomonidan qon aylanishidan chiqariladi. Xloromitronlarning singishiga o'xshash ularning so'rilishi retseptorlari tomonidan vositalangan va VLDL qoldiqlarida apoE mavjudligiga bog'liq (qo'shimcha ravishda 21-2, apoE va Altsgeymer kasalligi o'rtasidagi bog'liqlik tasvirlangan).
Shakl 21-40. Lipoproteinlar va lipidlar, va lipidlar transport orqali qon orqali lipoproteinlar ko'rinishida tashiladi, ular turli xil funktsiyalari va oqsil va lipidlarning turli xil tarkibiga ega bo'lgan bir nechta fraktsiyalarga birlashadi (21-1, 21-2-jadval) va bu fraktsiyalarning zichligiga to'g'ri keladi. Oziq-ovqat lipidlari chilomikronlarga birlashtiriladi, ularning tarkibidagi triatsilgliserollarning ko'pi lipoprotein lipazasi tomonidan adipoz va mushak to'qimalariga yuboriladi. Xylomikron qoldiqlari (asosan protein va xolesterolni o'z ichiga olgan) gepatotsitlar tomonidan ushlanadi. Jigardan endogen lipidlar va xolesterin yog'lar va mushak to'qimalariga VLDL shaklida etkaziladi. VLDLdan lipidlarning chiqishi (ba'zi apolipoproteinlarning yo'qolishi bilan birga) VLDLP ni asta-sekin LDL ga o'zgartiradi, bu xolesterolni ekstrahepatik to'qimalarga etkazib beradi yoki jigarga qaytaradi. Jigar VLDL, LDL qoldiqlarini va retseptorlar vositachilagan endotsitoz orqali chylomicronning qoldiqlarini ushlaydi. Ekstrahepatik to'qimalarda ortiqcha xolesterin LDL shaklida jigarga qaytariladi. Jigarda xolesterolning bir qismi safro tuzlariga aylanadi. b - ochlikdan keyin (chapda) va ko'p miqdordagi yog'li ovqatni iste'mol qilgandan so'ng olingan qon plazmasidan olingan namunalar (o'ngda). Yog'li oziq-ovqatlarni iste'mol qilish natijasida hosil bo'lgan ksilomikronlar plazma sutga tashqi o'xshashlikni beradi.

Triatsilgliserollarning yo'qolishi bilan VLDLning bir qismi VLDL qoldiqlariga aylantiriladi, shuningdek oraliq zichlikli lipoproteinlar (VLDL) deb ataladi, triatsilgliserollarni VLDLdan keyinchalik olib tashlashga imkon beradi. past zichlikli lipoproteinlar (LDL) (21-1-jadval). Xolesterin va xolesterin esterlariga juda boy bo'lgan, shuningdek apoB-100 bo'lgan LDL fraktsiyasi xolesterolni plazma membranalarida apoB-100 ni tan oladigan maxsus retseptorlarni olib yuradigan ekstrahepatik to'qimalarga o'tkazadi. Ushbu retseptorlar xolesterin va xolesterin esterlarini olishda vositachilik qiladi (quyida tavsiflangan).
Qo'shimcha 21-2.ApoE allellari Altsgeymer kasalligi tarqalishini aniqlaydi
Aholi populyatsiyasida apoE allellaridan iborat genlarni kodlaydigan uchta ma'lum variant (uchta allel) mavjud, apoE allellaridan APOEZ alleli odamlarda eng keng tarqalgan (taxminan 78%), APOE4 va APOE2 allellari mos ravishda 15 va 7% ni tashkil qiladi. APOE4 alleli Altsgeymer kasalligi bilan og'rigan odamlarga xosdir va bu munosabatlar kasallikning paydo bo'lish ehtimolini yuqori darajada taxmin qilish imkonini beradi. APOE4-ni meros qilib olgan odamlar kech Altsgeymer kasalligi rivojlanish xavfi yuqori. APOE4 uchun gomozigotli odamlar ushbu kasallikka nisbatan 16 baravar ko'proq, kasal bo'lganlarning o'rtacha yoshi 70 yoshni tashkil etadi. AROEZ ning ikki nusxasini meros qilib olgan odamlar uchun, aksincha, Altsgeymer kasalligining o'rtacha yoshi 90 yoshdan oshadi.
ApoE4 va Altsgeymer kasalligi o'rtasidagi bog'lanish uchun molekulyar asos hali ham noma'lum. Bundan tashqari, apoE4 Altsgeymer kasalligining asosiy sababi bo'lgan amiloid simlarining o'sishiga qanday ta'sir qilishi hali ham noma'lum (4-31-rasm, 1-rasmga qarang). Taxminlar apoE ning neyronlarning sitoskeletlari tarkibini barqarorlashtirishdagi mumkin bo'lgan roliga qaratilgan. ApoE2 va apoEZ oqsillari neyronlarning mikrotubulalari bilan bog'liq bo'lgan bir qator oqsillarga bog'lanadi, apoE4 esa bog'lamaydi. Bu neyronlarning o'limini tezlashtirishi mumkin. Ushbu mexanizm nima bo'lishidan qat'i nazar, ushbu kuzatuvlar apolipoproteinlarning biologik funktsiyalari to'g'risida tushunchamizni kengaytirishga umid beradi.
Lipoproteinlarning to'rtinchi turi - yuqori zichlikli lipoproteinlar (HDL), bu fraktsiya jigarda va ingichka ichakda nisbatan oz miqdordagi xolesterolni o'z ichiga olgan va xolesterin esterlaridan mutlaqo xoli bo'lgan oqsilga boy mayda zarralar shaklida hosil bo'ladi (21-40-rasm). HDL fraktsiyasida apoA-I, apoC-I, apoC-II va boshqa apolipoproteinlar mavjud (21-2-jadval), shuningdek lesitin-xolesterol-asiltransferaza (LC AT), bu lesitin (fosfatidilkolin) va xolesteroldan xolesterin esterlarini hosil bo'lishini katalizlaydi (21-41-rasm). Yangi hosil bo'lgan HDL zarralari yuzasida L CAT, xloromron xolesterol va fosfatidilkolin va VLDL qoldiqlarini xolesterin esterlariga aylantiradi, ular yadro hosil qila boshlaydi, yangi hosil bo'lgan diskoid HDL zarralarini etuk sferik HDL zarralariga aylantiradi. Ushbu xolesterolga boy lipoprotein jigarga qaytariladi, bu erda xolesterin "ajralib chiqadi", bu xolesterinning bir qismi safro tuzlariga aylanadi.
Shakl 21-41. Letsitin-xolesterin-asiltransferaza (L CAT) bilan reaktsiya katalizlanadi. Ushbu ferment HDL zarralari yuzasida mavjud va apoA-1 (HDL fraktsiyasining tarkibiy qismi) tomonidan faollashtiriladi. Xolesterin esterlari yangi hosil bo'lgan HDL zarralari ichida to'planib, ularni etuk HDLga aylantiradi.

HDL jigarda retseptorlari vositachilagan endotsitoz orqali so'rilishi mumkin, ammo kamida HDL xolesterolining bir qismi boshqa mexanizmlar orqali boshqa to'qimalarga etkaziladi. HDL zarralari jigar hujayralarining plazma membranasida va buyrak usti bezlari kabi steroidogen to'qimalarda SR - BI retseptorlari oqsillari bilan bog'lanishi mumkin. Ushbu retseptorlar endotsitozda vositachilik qilmaydi, ammo xolesterin va HDL fraktsiyasining boshqa lipidlarini hujayraga qisman va tanlab o'tkazish. HDL fraktsiyasi yana qonga kiradi, u erda xilomikron va VLDL qoldiqlaridan lipidlarning yangi qismlari kiradi. Xuddi shu HDL, shuningdek, ekstrahepatik to'qimalarda saqlanadigan xolesterolni ushlab, uni jigarga o'tkazishi mumkin teskari xolesterolni tashish (21-40-rasm). Reversiv transport variantlaridan birida, hosil bo'lgan HDLning xolesteringa boy hujayralardagi SR-BI retseptorlari bilan o'zaro ta'siri xolesterolni xujayralar sirtidan HDL zarralariga passiv tarqalishiga olib keladi va keyinchalik xolesterolni jigarga qaytaradi. Boy xolesterol hujayrasida teskari transportning yana bir variantida, HDL ajralganidan so'ng, apoA-I faol tashuvchisi, ABC oqsil bilan o'zaro ta'sir qiladi. ApoA-I (va ehtimol HDL) endotsitoz tomonidan so'riladi, so'ngra yana chiqarilib, xolesterin bilan to'ldirilib, jigarga etkaziladi.
Protein ABC1 ko'plab dorilarni tashuvchi katta oilaning tarkibiy qismidir, ba'zida bu tashuvchilar ABC tashuvchisi deb ataladi, chunki ularning barchasida ATP-ulanadigan kasetalar mavjud (ATP - bog'laydigan kasetalar), shuningdek, oltita transmembran spiralli ikkita transmembranli domenga egalar (bobga qarang). 11, v. 1). Ushbu oqsillar ko'plab ionlarni, aminokislotalarni, vitaminlarni, steroid gormonlarni va safro tuzlarini plazma membranalari orqali faol ravishda o'tkazadilar. Ushbu tashuvchilar oilasining yana bir vakili - bu CFTR oqsilidir, bu tsistoz fibroz bilan shikastlangan (qo'shimcha 11-3, v. 1).
Xolesterol esterlari hujayraga retseptorlar vositachilagan endotsitoz orqali kiradi
Qon oqimidagi har bir LDL zarracha apoB-100 ni o'z ichiga oladi, bu ma'lum sirt retseptorlari oqsillari tomonidan tan olingan -LDL retseptorlari xolesterolni ushlab turishi kerak bo'lgan hujayralar membranasida. LDL ning LDL retseptorlari bilan bog'lanishi endotsitozni keltirib chiqaradi, buning natijasida LDL va uning retseptorlari endosoma ichidagi hujayraga o'tadi (21-42-rasm). Oxir oqibatda endosoma xolesterin esterlarini gidroliz qiladigan, xolesterin va yog 'kislotalarini sitozolga chiqaradigan fermentlarni o'z ichiga olgan lizosoma bilan birlashadi. LDL dan olingan ApoB-100, shuningdek, sitozolda ajralib chiqadigan aminokislotalarni hosil qilish uchun parchalanadi, ammo LDL retseptorlari buzilishni oldini oladi va LDL olishda yana ishtirok etish uchun hujayra yuzasiga qaytadi. ApoB-100 VLDL-da ham mavjud, ammo uning retseptorlari bilan bog'laydigan domen LDL retseptorlari bilan bog'lana olmaydi, VLDLP-ni LDL-ga aylantirish retseptorlarni bog'laydigan domenni apoB-100-ga aylantiradi. Ushbu qon xolesterolini tashish yo'li va uning maqsadli to'qimalarda retseptorlari vositachilagan endotsitoz Maykl Braun va Jozef Goldshteyn tomonidan o'rganilgan.
Maykl Braun va Jozef Goldshteyn

Shakl 21-42. Xolesterolni retseptorlar vositasida endotsitoz bilan ushlash.

Bu tarzda hujayralarga kiradigan xolesterolni membrana ichiga kiritish yoki ACAT tomonidan qayta esterifikatsiya qilish mumkin (21-38-rasm) sitosol ichida lipid tomchilarida saqlash uchun. Qonning LDL fraktsiyasida etarli miqdordagi xolesterin mavjud bo'lganda, hujayra ichidagi xolesterolning to'planishi uning sintezi tezligini pasaytiradi.
LDL retseptorlari apoE bilan bog'lanadi va jigarda chylomicron va VLDL qoldiqlarini to'plashda muhim rol o'ynaydi. Ammo, agar LDL retseptorlari mavjud bo'lmasa (masalan, yo'qolgan LDL retseptorlari geni bilan sichqonchani shtammida), VLDL qoldiqlari va xloromronlar jigar tomonidan so'riladi, ammo LDL so'rilmaydi. Bu VLDL va chilomikron qoldiqlarining retseptorlari bilan ishlaydigan endotsitoz uchun yordamchi zaxira tizimining mavjudligini ko'rsatadi. Zaxira retseptorlaridan biri apoE va boshqa qator ligandlar bilan bog'laydigan lipoprotein retseptorlari bilan bog'liq bo'lgan LRP oqsilidir (lipoprotein retseptorlari bilan bog'liq protein).
Xolesterol biosintezining bir necha darajalari
Xolesterol sintezi murakkab va baquvvat jarayondir, shuning uchun tanada xolesterin biosintezini tartibga solish mexanizmi mavjudligi aniq, bu oziq-ovqat bilan birga qo'shimcha ravishda uning miqdorini to'ldiradi. Sutemizuvchilarda xolesterin ishlab chiqarish hujayra ichidagi kontsentratsiya bilan boshqariladi
xolesterol va gormonlar glyukagon va insulin. HMG - CoA ni mevalonatga o'tkazish bosqichi (21-34-rasm) xolesterin hosil bo'lishining metabolik yo'lidagi tezlikni cheklaydi (tartibga solishning asosiy nuqtasi). Ushbu reaktsiya HMG - CoA reduktaza bilan katalizlanadi. Xolesterin darajasining o'zgarishiga javoban tartibga solish, HMG - CoA reduktazasini kodlovchi genni transkriptsiyalashning oqlangan tizimi bilan ta'minlanadi. Ushbu gen, xolesterolni va to'yinmagan yog'li kislotalarni so'rib olish va sintez qilishda ishtirok etadigan 20 dan ortiq boshqa fermentlar bilan birgalikda oqsil hosil bo'lishining sterol-tartibga soluvchi elementi (SREBP, sterolni tartibga soluvchi elementlar bilan bog'laydigan oqsillar) deb ataladigan oqsillar deb ataladigan kichik bir oila tomonidan boshqariladi. . Sintezdan so'ng, bu oqsillar endoplazmatik retikulumga kiritiladi. Faqatgina eriydigan SREBP amino terminali Ch da tasvirlangan mexanizmlardan foydalangan holda transkripsiya faollashtiruvchisi vazifasini bajaradi. 28 (3-oyat). Biroq, bu domen yadroga kirish huquqiga ega emas va SREBP molekulasida qolganda genni faollashtirishda qatnasha olmaydi. HMG geni - CoA reduktazasi va boshqa genlarning transkripsiyasini faollashtirish uchun transkriptiv faol domen SREBP ning qolgan qismidan proteolitik bo'linish bilan ajratilgan. Xolesterin miqdori ko'p bo'lganida SREBP oqsillari faol emas va ERAPga boshqa bir protein (SREBP - ajratuvchi faollashtiruvchi oqsil) bilan biriktirilgan (21-43-rasm). Bu sterol sensori sifatida ishlaydigan xolesterolni va boshqa bir qator sterollarni bog'laydigan SCAP. Sterol darajasi yuqori bo'lganda, SCAP - SREBP kompleksi, ehtimol, butun kompleksni ER da saqlaydigan boshqa protein bilan o'zaro ta'sir qiladi. Hujayra ichidagi sterollarning miqdori pasayganda, SCAP tarkibidagi muvofiqlik o'zgarishi tutilish faolligini yo'qotishiga olib keladi va SCAP - SREBP kompleksi vesikullar ichidan Golgi kompleksiga ko'chadi. Golgi majmuasida SREBP oqsillari ikki xil proteazadan ikki marta ajratiladi, ikkinchi ajralish amino-terminal domenini sitozolga chiqaradi. Ushbu domen yadroga o'tadi va maqsadli genlarning transkripsiyasini faollashtiradi. SREBP aminokislotasi domeni qisqa umrga ega va proteasomalar tomonidan tez buziladi (27-48-rasm, 3-rasmga qarang). Sterol darajasi etarlicha ko'tarilganda, aminokislotalar bilan SR EBP oqsil domenlarining proteolitik chiqishi yana bloklanadi va mavjud faol domenlarning proteazom degradatsiyasi maqsadli genlarning tezda yopilishiga olib keladi.
Shakl 21-43. SR EBP ni faollashtirish. Sterol bilan boshqariladigan SREB P oqsillari (yashil rang) sintezdan so'ng darhol ERga kiritilib, S CAP (qizil rang) bilan kompleks hosil qiladi. (N va C oqsillarning amin va karboksil uchlarini bildiradi.) S-CAP bilan bog'langan holatda SRE BP oqsillari faol emas. Sterol darajasi pasayganda, SR EBP-S CAP kompleksi Golgi kompleksiga o'tadi va SR EBP oqsillari ketma-ket ikki xil proteaz bilan ajralib chiqadi. Ozod qilingan aminokislotalar terminali SR EBP oqsil domeni yadroga ko'chib o'tadi, u erda sterol bilan boshqariladigan genlarning transkripsiyasini faollashtiradi.

Xolesterol sintezi boshqa bir qancha mexanizmlar tomonidan ham tartibga solinadi (21-44-rasm). Gormonal nazorat NM G-CoA reduktazasining kovalent modifikatsiyasi bilan vositachilik qilinadi. Ushbu ferment fosforilatlangan (nofaol) va deposforillangan (faol) shakllarda mavjud. Glyukagon fermentning fosforlanishini (inaktivatsiyasini) rag'batlantiradi, insulin esa fermentni faollashtiradigan va xolesterin sintezini yoqadigan dehosforilatsiyani rag'batlantiradi. Xolesterolning yuqori hujayra ichidagi kontsentratsiyasi ASAT ni faollashtiradi, bu esa xolesterolni cho'ktirish uchun esterifikatsiyasini oshiradi. Va nihoyat, hujayrali xolesterolning yuqori miqdori LDL retseptorlarini kodlaydigan genning transkripsiyasini inhibe qiladi, bu retseptorning ishlab chiqarilishini va shuning uchun qondan xolesterolni olishni kamaytiradi.
Shakl 21-44. Xolesterin miqdorini boshqarish xolesterolni oziq-ovqatdan sintezi va so'rilishi o'rtasidagi muvozanatni ta'minlaydi. Glyukagon NM G -CoA reduktsazining fosforlanishini (inaktivatsiyasini) osonlashtiradi, insulin esa defosforlanishni (faollashishni) ta'minlaydi. X - NM G -CoA reduktazasining proteolizini rag'batlantiradigan aniqlanmagan xolesterol metabolitlari.

Nazorat qilinmagan xolesterin odamlarda jiddiy kasalliklarga olib kelishi mumkin. Sintezlangan xolesterin va oziq-ovqatdan olingan xolesterin miqdori membranani yig'ish, safro tuzlari va steroidlarni sintez qilish uchun zarur bo'lgan miqdordan oshib ketganda, qon tomirlarida xolesterinning patologik to'planishi (aterosklerotik plaklar) paydo bo'lishi mumkin, bu ularning tiqilib qolishiga olib keladi (ateroskleroz). Sanoati rivojlangan mamlakatlarda bu o'limning asosiy sababi bo'lgan koronar arteriyalarning obstruktsiyasi tufayli yurak etishmovchiligi. Aterosklerozning rivojlanishi qonda yuqori xolesterin va ayniqsa LDL fraktsiyasi bilan yuqori darajadagi xolesterin miqdori bilan bog'liq va HDL qonining yuqori darajasi, aksincha, qon tomirlarining holatiga ijobiy ta'sir qiladi.
Irsiy giperkolesterolemiya (irsiy nuqson) bilan qonda xolesterin darajasi juda yuqori - bu odamlarda bolalikdan qattiq ateroskleroz rivojlanadi. LDL nuqsonli retseptorlari tufayli LDL xolesterolini etarli darajada retseptorlari vositasida qabul qilish sodir bo'ladi. Natijada, xolesterin qon oqimidan chiqarilmaydi, u to'planib, aterosklerotik blyashka shakllanishiga hissa qo'shadi. Qonda ortiqcha xolesterolga qaramay, endogen xolesterol sintezi davom etmoqda, chunki hujayradan tashqari xolesterin hujayralararo sintezni tartibga solish uchun hujayraga kira olmaydi (21 -44-rasm).Irsiy giperkolesterolemiya va sarum xolesterolining ko'tarilishi bilan bog'liq boshqa kasalliklarni davolashda statin sinflari qo'llaniladi. Ulardan ba'zilari tabiiy manbalardan olinadi, boshqalari farmatsevtika sanoati tomonidan sintez qilinadi. Statinlar mevalonatga o'xshaydi (qo'shimcha 21-3) va NMS-CoA reduktazasining raqobatbardosh inhibitori hisoblanadi.
21-3-ilova. TIBBIYOT. Lipit gipotezasi va statinlarning yaratilishi
Rivojlangan mamlakatlarda yurak tomirlari kasalligi (CHD) o'limning asosiy sababidir. Yurakka qon olib keladigan koronar arteriyalarning torayishi aterosklerotik plaklar deb ataladigan yog 'birikmalarining paydo bo'lishi natijasida yuzaga keladi, bu plitalarda xolesterin, fibrillalar oqsillari, kaltsiy, trombotsitlar va hujayralar parchalari mavjud. XX asrda. Arterial obstruktsiya (ateroskleroz) va qon xolesterolining o'zaro bog'liqligi haqida faol munozaralar bo'lib o'tdi. Ushbu muhokamalar va ushbu yo'nalishdagi faol tadqiqotlar xolesterolni pasaytiradigan samarali dorilarni yaratishga olib keldi.
1913 yilda taniqli rus olimi va eksperimental patologiya sohasi mutaxassisi N.N. Anichkov "Xolesterolga boy oziq-ovqat bilan oziqlangan quyonlar keksa odamlarning tomirlarida aterosklerotik blyashka o'xshash qon tomirlariga zarar etkazishini ko'rsatdi" deb nomlangan asarini nashr etdi. Anichkov bir necha o'n yillar davomida o'z tadqiqotlarini olib bordi va natijalarini taniqli G'arb jurnallarida nashr etdi. Afsuski, uning ma'lumotlari odamlarda ateroskleroz rivojlanishining modeli uchun asos bo'lmadi, chunki o'sha paytda bu kasallik qarishning tabiiy natijasi ekanligi haqidagi gipoteza mavjud edi va uni oldini olish mumkin emas. Ammo, zardobdagi xolesterin va aterosklerozning rivojlanishi (lipid gipotezasi) o'rtasidagi aloqani asta-sekin to'plash va 1960 yillarda. ba'zi tadqiqotchilar ushbu kasallikni dori-darmonlar bilan davolash mumkinligini aniq aytishdi. Biroq, 1984 yilda AQSh Milliy sog'liqni saqlash instituti tomonidan o'tkazilgan xolesterolning rolini keng o'rganish natijalari (koronar boshlang'ich profilaktika sinovi) nashr etilgunga qadar qarama-qarshi nuqtai nazar mavjud edi. Miyokard infarkti va qon xolesterinining pasayishi bilan insult chastotasining statistik jihatdan sezilarli darajada pasayishi ko'rsatilgan. Ushbu tadqiqotda xolesterolni pasaytirish uchun safro kislotalarini bog'laydigan anion almashinadigan qatronlar bo'lgan xolesterin ishlatilgan. Natijalar yangi, yanada kuchli terapevtik dori-darmonlarni izlashni rag'batlantirdi. Aytishim kerakki, ilmiy dunyoda lipid gipotezasining haqiqiyligiga shubha faqat 1980 yillarning oxiri - 90-yillarning boshlarida statinlar paydo bo'lishi bilan butunlay yo'q bo'lib ketdi.
Birinchi statinni Akira Endo Tokiodagi Sankyoda topdi. Endo o'z asarini 1976 yilda nashr etgan, garchi u bir necha yil davomida xolesterin metabolizmi muammosi bilan shug'ullangan. 1971 yilda u o'sha davrda o'rganilgan antibiotiklarni ishlab chiqaradigan qo'ziqorin ishlab chiqaruvchilarida xolesterin sintezi inhibitörleri ham bo'lishi mumkinligini taklif qildi. Bir necha yillik intensiv mehnat davomida u ijobiy natijaga erishgunga qadar turli xil qo'ziqorinlarning 6000 dan ortiq madaniyatini tahlil qildi. Olingan birikma kompaktin deb nomlandi. Ushbu modda itlar va maymunlarda xolesterolni kamaytirdi. Ushbu tadqiqotlar Maykl Braun va Jozef Goldshteynning Texas janubi-g'arbiy tibbiyot maktabidagi universiteti e'tiborini tortdi. Braun va Goldshteyn, Endo bilan birgalikda, birgalikda o'rganishni boshlashdi va uning ma'lumotlarini tasdiqlashdi. Birinchi klinik sinovlarning asosiy yutuqlari farmatsevtika kompaniyalarining ushbu yangi dori vositalarini ishlab chiqishda ishtirok etishidir. Merckda Alfred Alberts va Roy Vagelos boshchiligidagi guruh qo'ziqorin madaniyati bo'yicha yangi skriningni boshladilar va jami 18 ta madaniyatni tahlil qilish natijasida yana bir faol dorini topdilar. Yangi modda lovastatin deb ataladi. Ammo, shu bilan birga, kompaktinning yuqori dozalarini itlarga yuborish 1980-yillarda saraton kasalligini rivojlanishiga va yangi statinlarni qidirishga olib keladi degan fikr keng tarqalgan edi. to'xtatildi. Ammo, bu vaqtga kelib, oilaviy giperkolesterolemiya bilan og'rigan bemorlarni davolash uchun statinlardan foydalanishning foydalari allaqachon ayon bo'ldi. Xalqaro ekspertlar va Oziq-ovqat va farmatsevtika idorasi (FDA, AQSh) bilan o'tkazilgan ko'plab maslahatlashuvlardan so'ng Merck lovastatinni ishlab chiqara boshladi. Keyingi yigirma yil ichida olib borilgan keng qamrovli tadqiqotlar lovastatinning kanserogen ta'sirini va undan keyin paydo bo'lgan yangi dorilarni aniqlamadi.
Shakl 1. Statinlar NM G-CoA reduktazasining ingibitorlaridir. NM G -CoA reduktazasining ta'sirini kamaytiradigan mevalonat va to'rtta farmatsevtika mahsulotlarining (statinlarning) tuzilishini taqqoslash.

Statinlar HMG - CoA - reduktaza ta'sirini inhibe qiladi, mevalonatning tuzilishini taqqoslaydi va shu bilan xolesterin sintezini bloklaydi. LDL retseptorlari genining bir nusxasidagi nuqson tufayli kelib chiqqan giperkolesterolemiya bilan og'rigan bemorlarda, lovastatinni qabul qilganda, xolesterin miqdori 30 foizga kamayadi. Preparat safro kislotalarini bog'laydigan va ichaklardan teskari so'rilishini oldini oluvchi maxsus qatronlar bilan birgalikda samaraliroq hisoblanadi.
Hozirgi vaqtda statinlar qon plazmasidagi xolesterolni pasaytirish uchun ko'pincha ishlatiladi. Har qanday dori-darmonlarni qabul qilganda, ularning nojo'ya ta'sirlari haqida savol tug'iladi. Ammo, statinlar bo'lsa, aksariyat yon ta'siri, aksincha, ijobiydir. Ushbu dorilar qon oqimini rag'batlantirishi, allaqachon mavjud bo'lgan aterosklerotik plaklarni mahkamlashi (qon tomirlari devorlaridan ajralmasligi va qon oqimiga xalaqit bermasligi uchun), trombotsitlar yig'ilishini inhibe qilishi, shuningdek qon tomirlari devorlarida yallig'lanish jarayonlarini susaytirishi mumkin. Birinchi marta statinlarni qabul qilgan bemorlarda bu ta'sir xolesterin darajasi pasayishidan oldin ham namoyon bo'ladi va, ehtimol, izoprenoid sintezining susayishi bilan bog'liq. Albatta, statinlarning har qanday yon ta'siri ham foydali emas. Ba'zi bemorlarda (odatda statinlarni qabul qiladiganlar orasida xolesterolni kamaytiradigan boshqa dorilar bilan birgalikda) mushaklardagi og'riq va mushaklarning zaifligi, ba'zida esa ancha kuchli shaklda bo'lishi mumkin. Statinlarning boshqa juda ko'p yon ta'siri ham qayd etilgan, ular xayriyatki kamdan-kam hollarda uchraydi. Bemorlarning ko'pchiligida statinlarni qabul qilish yurak-qon tomir kasalliklarining rivojlanishiga to'sqinlik qilishi mumkin. Boshqa har qanday dori-darmonlar singari, statinlar faqat shifokor tomonidan tavsiya etilganidek qo'llanilishi kerak.

HDL xolesterolining irsiy yo'qligida, xolesterin darajasi juda past, Tanger kasalligi bilan, xolesterin deyarli aniqlanmaydi. Ikkala genetik kasallik ham ABC1 oqsilidagi mutatsiyalar natijasida yuzaga keladi. HDL-bo'sh xolesterol fraktsiyasi ABC1 etishmovchiligidagi hujayralardagi xolesterolni ushlab turolmaydi va xolesterinni yo'qotgan hujayralar qondan tezda chiqarilib, yo'q qilinadi. HDL va Tanger kasalligining irsiy yo'qligi juda kam uchraydi (tanga kasalligi bilan yuzdan kam oila dunyoga ma'lum), ammo bu kasalliklar HDL plazma darajasini boshqarishda ABC1 oqsilining rolini namoyish etadi. Past plazma HDL darajasi koronar arteriya shikastlanishining yuqori darajasi bilan bog'liq bo'lganligi sababli, ABC1 oqsil HDL darajasini tartibga solish uchun mo'ljallangan dorilar uchun foydali bo'lishi mumkin. ■
Steroid gormonlar xolesterol va uning oksidlanishining yon zanjirini ajratish orqali hosil bo'ladi.
Biror kishi barcha steroid gormonlarini xolesteroldan oladi (21-45-rasm). Adrenal korteksda ikkita steroid gormon sintezlanadi: mineralkortikoidlar,noorganik ionlarning (Na +, C l - va HC O) so'rilishini tartibga soluvchi 3 -) buyraklarda va glyukokortikoidlar, bu glyukoneogenezni tartibga solishga va yallig'lanish reaktsiyasini kamaytirishga yordam beradi. Jinsiy gormonlar erkaklar va ayollarning jinsiy hujayralarida va yo'ldoshda ishlab chiqariladi. Ular orasida progesteron ayollarning reproduktiv tsiklini boshqaruvchi androgenlar (masalan, testosteron) va estrogenlar (estradiol), o'z navbatida erkaklar va ayollarda ikkilamchi jinsiy xususiyatlarning rivojlanishiga ta'sir qiladi. Steroid gormonlari juda past konsentratsiyalarda ta'sir qiladi va shuning uchun nisbatan oz miqdorda sintezlanadi. Safro tuzlari bilan solishtirganda, steroid gormonlarini ishlab chiqarish uchun nisbatan oz miqdordagi xolesterin iste'mol qilinadi.

Shakl 21-45. Ba'zi steroid gormonlar xolesteroldan hosil bo'ladi. Ushbu birikmalarning ayrimlarining tuzilishi sek. 10-19, 1-bet.

Steroid gormonlar sintezi xolesterinning C-17 D-halqasining "yon zanjirida" bir nechta yoki barchasini uglerod atomlarini yo'q qilishni talab qiladi. Yon zanjirni olib tashlash steroidogen to'qimalarning mitoxondriyalarida sodir bo'ladi. Olib tashlash jarayoni yonma-yon joylashgan ikkita qo'shni uglerod atomining gidroksillanishi (C-20 va C-22), so'ngra ular orasidagi bog'lanishning yorilishidan iborat (21-46-rasm). Turli xil gormonlar hosil bo'lishi kislorod atomlarini kiritishni ham o'z ichiga oladi. Steroid biosintezi paytida barcha gidroksillanish va oksidlanish reaktsiyalari NA D PH, O dan foydalanadigan aralash funktsional oksidazalar (21-1 qo'shimcha) bilan katalizlanadi. 2 va P-450 mitoxondrial sitoxrom.
Shakl 21-46. Steroid gormonlar sintezida yon zanjirning yorilishi. Qo'shni uglerod atomlarini oksidlovchi aralash funktsiyaga ega bo'lgan bu oksidaza tizimida P-450 sitoxromi elektron tashuvchisi sifatida ishlaydi. Bu jarayonda elektron tashiydigan oqsillar, adrenodoksin va adrenodoksin reduktaza ham ishtirok etadi. Yon zanjirni ajratish tizimi adrenal korteksning mitoxondriyalarida topilgan, bu erda steroid faol ishlab chiqariladi. Pregnenolon boshqa barcha steroid gormonlarning ta'sirchan qismidir (21-45-rasm).

Xolesterol biosintezi vositachilari boshqa ko'plab metabolik yo'llarda qatnashadilar.
Xolesterol biosintezi vositachisi rolidan tashqari, izopentenil pirosfat turli xil biologik funktsiyalarni bajaradigan juda ko'p sonli biomolekulalarni sintez qilishda faollashtirilgan prekursor bo'lib xizmat qiladi (21-47-rasm). Bularga A, E va K vitaminlari, karotin kabi o'simlik pigmentlari va xlorofill fitol zanjiri, tabiiy kauchuk, ko'plab efir moylari (masalan, limon yog'ining xushbo'y asosi, evkalipt, mushk), metamorfozni tartibga soluvchi balog'atga etmagan gormon, dolichollar kiradi. mitoxondriyalar va xloroplastlarda polisakkaridlar, ubikinon va plastokinon - elektron tashuvchilarning kompleks sintezida lipid erituvchi tashuvchilar bo'lib xizmat qiladi. Bu barcha molekulalar tarkibidagi izoprenoidlardir. Tabiatda 20 mingdan ortiq turli xil izoprenoidlar topilgan va har yili yuzlab yangilari haqida xabar beriladi.
Shakl 21-47. Izoprenoidlarning biosintezi haqida umumiy rasm. Bu erda keltirilgan ko'plab mahsulotlarning tuzilishi bobda keltirilgan. 10 (1-oyat).

Prenilatsiya (izoprenoidning kovalent biriktirilishi, 27-35-rasmga qarang) sutemizuvchi hujayralar membranalarining ichki yuzasida oqsillarni langar qiladigan keng tarqalgan mexanizmdir (11-14-rasmlarga qarang). Ba'zi oqsillarda bog'langan lipid 15 karbonli farnesil guruhi, boshqalarida bu 20 karbonli geranil geranil guruhi. Ushbu ikki turdagi lipidlar turli xil fermentlarni biriktiradilar. Prenilatsiya reaktsiyalari oqsillarni turli lipidlarga biriktirilganiga qarab turli membranalarga yo'naltirishi mumkin. Proteinni ko'paytirish izopren hosilalari - xolesterol metabolik yo'li ishtirokchilari uchun yana bir muhim rol o'ynaydi.
21.4-qismning qisqacha mazmuni Xolesterin, Ukol va Izoprenoidlarning biosintezi
■ Xolesterin atsetil-CoA dan reaction-gidroksi-β-metilglyutaril-CoA, mevalonat, ikkita faollashtirilgan izopren dimetilally pirofosfat va izopentenil pirofosfat kabi vositachilar orqali murakkab reaktsiya ketma-ketligida hosil bo'ladi. Izopren birliklarining kondensatsiyasi tsiklik bo'lmagan skalenni beradi, u kondensatsiyalangan halqali tizim va steroid yon zanjirini hosil qiladi.
■ Xolesterol sintezi gormonal nazorat ostida va qo'shimcha ravishda kovalent modifikatsiya va transkripsiyani tartibga solish natijasida yuzaga keladigan hujayra ichidagi xolesterol kontsentratsiyasining ortishiga to'sqinlik qiladi.
■ Xolesterin va xolesterol esterlari qonda plazma lipoproteinlari sifatida tashiladi. VLDL fraktsiyasi xolesterolni, xolesterin esterlarini va triatsilgliserollarni jigardan boshqa to'qimalarga o'tkazadi, bu erda triatsilgliserollarni lipoprotein lipazasi bilan yopishadi va VLDL LDLga aylanadi. Xolesterin va xolesterin esterlarida boyitilgan LDL fraktsiyasi bilvosita endotsitoz bilan retseptorlari tomonidan ushlanadi, LDL tarkibidagi B-100 apolipoprotein esa plazma membranasi retseptorlari tomonidan tan olinadi. HDL qondan xolesterolni olib tashlaydi va uni jigarga o'tkazadi. Oziqlanish sharoitlari yoki xolesterol metabolizmidagi irsiy nuqsonlar ateroskleroz va miyokard infarktiga olib kelishi mumkin.
■ Steroid gormonlar (glyukokortikoidlar, mineralokortikoidlar va jinsiy gormonlar) xolesteroldan yon zanjirni almashtirish va kislorod atomlarini uzukning steroid tizimiga kiritish orqali hosil bo'ladi. Boshqa ko'plab izoprenoid birikmalari xolesterol bilan birga izopentenil pirosfat va dimetilillil pirofosfat kondensatsiyasi orqali mevalonatdan hosil bo'ladi.
■ Ba'zi bir oqsillarni profilaktikasi ularni hujayra membranalari bilan bog'lanadigan joylarga yo'naltiradi va ularning biologik faolligi uchun muhimdir.
Savol 48. Yuqori yog'li kislotalarning metabolizmini tartibga solish (β-oksidlanish va biosintez). Malonil CoA sintezi. Atsetil CoA karboksilaza, uning faoliyatini tartibga solish. Akil Co-A ni mitoxondriyaning ichki membranasi orqali tashish.
Asosiy
fenilalanin miqdori iste'mol qilinadi
2 usulda:
yoqiladi
sincaplarda,
buriladi
tirozinda.
Burish
fenilalanin birinchi navbatda tirozinga
ortiqcha olib tashlash uchun zarur
fenilalanin, chunki yuqori konsentratsiya
Uning hujayralar uchun zaharli. Ta'lim
tirozin aslida muhim emas
chunki bu aminokislotaning etishmasligi
hujayralarda deyarli bunday bo'lmaydi.
Asosiy
fenilalanin metabolizmi boshlanadi
gidroksillanishi bilan (9-29-rasm),
natijada tirozin paydo bo'ladi.
Ushbu reaktsiya ma'lum bir tomonidan katalizlanadi
monoksi-nase - fenilalanin gidrasi (zsilaza,
bu koferment vazifasini bajaradi
tetrahidrobiopterin (N4BP).
Ferment faolligi ham bog'liq
Fe2 mavjudligi.
Ichida
jigar birinchi navbatda tezlashtirilgan mobilizatsiya
glikogen (7-bo'limga qarang). Biroq, aktsiyalar
Jigarda glikogen tugaydi
18-24 soatlik ro'za. Asosiy manba
zaxiralari tugashi bilan glyukoza
glikogen glyukoneogenezga aylanadi,
orqali tezlasha boshlaydi
Shakl
11-29. Asosiy metabolik o'zgarishlar
changni yutish moslamasini o'zgartirganda energiya
postabsorbent holat. KT
- keton tanalari, FA - yog 'kislotalari.
4-6 soat
oxirgi ovqatdan keyin. Substratlar
glitserin glyukoza sintezi uchun ishlatiladi,
aminokislotalar va laktat. Yuqori
glyukagon kontsentratsiyasi sintezi darajasi
tufayli yog 'kislotalari kamayadi
fosforillash va inaktivatsiya
atsetil CoA karboksilaza va nisbati
p-oksidlanish kuchayadi. Ammo,
jigarga yog 'etkazib berishning ko'payishi
tashigan kislotalar
yog'li omborlardan. Asetil-KoA hosil bo'lgan
yog 'kislotalarini oksidlanishida ishlatiladi
jigarda keton tanalarini sintez qilish uchun.
Ichida
konsentratsiyasining ortishi bilan yog 'to'qimasi
glyukagon sintez tezligini pasaytirdi
TAG va lipoliz rag'batlantiriladi. Rag'batlantirish
lipoliz - faollashuv natijasi
gormonga sezgir TAG lipaz
glyukagon ta'siri ostida adipotsitlar.
Yog 'kislotalari muhim ahamiyatga ega
jigar, mushaklar va energiya manbalari
yog 'to'qimasi.
Shunday qilib
Shunday qilib, postabsorbtsiya davrida
qonda glyukoza kontsentratsiyasi saqlanib qoladi
80-100 mg / dl va yog 'darajasida
kislotalar va keton tanalari ko'payadi.
Shakar
diabet - bu kasallik
mutlaq yoki nisbiy tufayli
insulin etishmovchiligi.
A
Shakarning asosiy klinik shakllari
diabet
Ko'ra
Jahon tashkiloti
sog'liqni saqlash diabet
farqlarga ko'ra tasniflanadi
genetik omillar va klinik
ikkita asosiy shakl: diabet
I tur - insulinga bog'liq (IDDM) va diabet
II toifa - insulinsiz mustaqil (NIDDM).
Tartibga solish
regulyatsion ferment sintezi
lcd - asetil CoA karboksilaza sintezi.
Ushbu ferment bir necha tomonidan tartibga solinadi
yo'llari.
Faollashtirish / ajratish
ferment subunit komplekslari. Ichida
atsetil CoA karboksilazasining faol bo'lmagan shakli
alohida komplekslarni ifodalaydi,
ularning har biri 4 qismdan iborat.
Fermentning faollashtiruvchisi sitratdir. Bu rag'batlantiradi
komplekslarning kombinatsiyasi, natijada
bu orqali ferment faolligi oshadi
. Inhibitor-palmitoyl-CoA. U qo'ng'iroq qiladi
murakkab dissosiatsiya va pasayish
ferment faolligi.
Fosforillash / Deposforillash
atsetil CoA karboksilaza. Ichida
postabsorbtsiya holati yoki
jismoniy ish glucagonized
adenilat siklazasi orqali adrenalin
tizim prokinaza A va orqali faollashadi
subunit fosforillanishini rag'batlantirish
atsetil CoA karboksilaza. Fosforitlangan
ferment faol emas va yog 'sintezi
kislotalar to'xtaydi.
Absorbent
insulin fosfatazani faollashtiradi,
va atsetil-CoA karboksilaza kiradi
dehosforlangan holat. Keyin
sitrat ta'siri ostida sodir bo'ladi
ferment protomerlarining polimerizatsiyasi va
u faollashadi. Faollashtirishga qo'shimcha ravishda
ferment, sitrat boshqasini amalga oshiradi
Suyuq kristall sintezidagi funksiya. Absorbent
jigar hujayralari mitoxondriyalaridagi davr
unda sitrat to'planadi, bunda
oksil qoldig'i tashiladi
sitosol.
Tartibga solish
β-oksidlanish darajasi.
Β-oksidlanish-metabolik yo'l,
CPE va umumiy ish bilan mustahkam bog'langan
katabolizm usullari. Shuning uchun uning tezligi
uchun zarur bo'lgan hujayra tomonidan boshqariladi
energiya ATP / ADP va NADH / NAD nisbati, shuningdek CPE va
katabolizmning umumiy yo'li. Tezlik
To'qimalarda β-oksidlanish mavjudligiga bog'liq
substrat, ya'ni.
yog 'miqdori to'g'risida
mitoxondriyaga kiradigan kislotalar.
Bepul yog'li kislota konsentratsiyasi
faollashgandan so'ng qonda ko'tariladi
ro'za paytida yog 'to'qimasida lipoliz
glyukagon ta'siri ostida va jismoniy paytida
adrenalin ta'siri ostida ishlash. Bularda
yog 'kislotalari aylanadi
ustun energiya manbai
natijada mushaklar va jigar uchun
β-oksidlanish NADH va atsetil-CoA inhibe qiladigan tomonidan hosil bo'ladi
piruvat dehidrogenaz kompleksi.
Piruvat hosil bo'lishining transformatsiyasi
glyukozadan atsetil-CoA ga o'tish jarayoni sekinlashadi.
Oraliq metabolitlar to'planadi
glikoliz va ayniqsa glyukoza-6-fosfat.
Glyukoza-6-fosfat geksokinazni inhibe qiladi
va shuning uchun tushkunlik
jarayonda glyukoza foydalanish
glikoliz. Shuning uchun, ustun
LCD-ni asosiy manba sifatida ishlatish
mushak to'qimasida va jigarda energiya
asab to'qimalari uchun glyukozani saqlaydi va
qizil qon hujayralari.
Β-oksidlanish darajasi
ferment faolligiga bog'liq
karnitin akriltransferazalari I.
Jigarda bu ferment inhibe qilinadi.
hosil bo'lgan molonil CoA
LCD ning biosintezi bilan Yutish davrida
jigarda glikoliz faollashadi va
atsetil-CoA hosil bo'lishi oshadi
piruvatdan. Birinchi sintez reaktsiyasi
asetil-CoA ning malonil-CoA ga lCD konversiyasi.
Malonil-KoA suyuq kristallning β oksidlanishini inhibe qiladi,
sintez uchun ishlatilishi mumkin
yog '.
Ta'lim
atsetil-CoA-tartibga soluvchi malonil-CoA
LCDdagi biosintezdagi reaktsiya. Birinchi reaktsiya
asetil-CoA ga lcd konvertatsiya qilish
malonil CoA. Katalitik ferment
bu reaktsiya (atsetil Coa karboksilaza),
ligazlar sinfiga tegishli. U o'z ichiga oladi
kovalent bog'langan biotin. Birinchisida
co2 kovalent reaktsiya bosqichlari
energiya tufayli biotinga bog'laydi
ATP, 2 bosqichda COO uzatildi
Atsetil-CoAda malonil-CoA hosil bo'ladi.
Asetil CoA karboksilaza fermenti faolligi
keyingi barcha tezlikni belgilaydi
sintez reaktsiyalari lc
sitrat sitozolda fermentni faollashtiradi
atsetil CoA karboksilaza. Malonyl CoA in
o'z navbatida oliyning o'tishiga xalaqit beradi
yog 'kislotalari sitozoldan matritsaga qadar
faoliyatni inhibe qiluvchi mitoxondriya
tashqi atsetil CoA: karnitin asiltransferaza,
yuqori oksidlanishni to'xtatadi
yog 'kislotalari.
Asetil-CoA Oksaloatsetat →
HS-CoA Citrate

HSCOA ATP Citrate → Asetil-CoA ADP Pi Oxaloatsetat
Asetil-CoA
sitoplazmada boshlang'ich substrat bo'lib xizmat qiladi
LCD va oksaloatsetatni sintez qilish uchun
tsitozol tarkibidagi o'zgarishlarga uchraydi
natijada piruvat hosil bo'ladi.
Xolesterin biosintezi
Xolesterol biosintezi endoplazmatik retikulumda uchraydi. Molekuladagi barcha uglerod atomlarining manbai yog 'kislotalari sintezidagi kabi sitratda mitoxondriyalardan kelib chiqqan atsetil-SCoA hisoblanadi. Xolesterin biosintezi 18 ATP molekulasini va 13 NADPH molekulasini iste'mol qiladi.
Xolesterolning shakllanishi 30 dan ortiq reaktsiyalarda uchraydi, ularni bir necha bosqichda guruhlash mumkin.
1. Mevalonik kislota sintezi.
Dastlabki ikkita sintez reaktsiyasi ketogenez reaktsiyalariga to'g'ri keladi, ammo 3-gidroksi-3-metilglyutaril-ScoA sintezidan keyin ferment fermentga kiradi. gidroksimetil-glutaril-ScoA reduktazasi (HMG-SCOA reduktaz) mevalon kislotasini hosil qiladi.
 |