Xolesterin sintezini tartibga solish
Xolesterin sintezini tartibga solish - uning asosiy fermenti (HMG-CoA reduktaza) turli yo'llar bilan amalga oshiriladi.
HMG reduktazasining fosforillanishi / deposforillanishi. Insulin / glyukagon nisbati oshishi bilan bu ferment fosforilatlanadi va faol holatga o'tadi. Insulinning ta'siri 2 ta ferment orqali amalga oshiriladi.
HMG-CoA redüktaz kinaz fosfataza, bu kinazani faol bo'lmagan defosforillangan holatga aylantiradi:
Fosfotaza HMG-CoA reduktazasini uni dehosforlangan faol holatga o'tkazish orqali. Ushbu reaktsiyalarning natijasi HMG-CoA reduktazasining dehosforlangan faol shaklini shakllantirishdir.
Shunday qilib, assimilyatsiya davrida xolesterin miqdori oshadi. Ushbu davrda xolesterin - asetil - CoA sintezi uchun dastlabki substratning mavjudligi oshadi (uglevodlar va yog'larni o'z ichiga olgan oziq-ovqatlarni iste'mol qilish natijasida, chunki CoA asetil glyukoza va yog 'kislotalari parchalanishi jarayonida hosil bo'ladi).
Postabsorbent holatda, proteingenaza A orqali glyukagon HMG - CoA - reduktsazning fosforlanishini rag'batlantiradi va uni faol bo'lmagan holatga aylantiradi. Ushbu harakat glyukagon bir vaqtning o'zida HMG-CoA reduktazasining fosforlanishini va inaktivatsiyasini rag'batlantiradi va shu bilan HMG-CoA redüktazasini fosforillangan faol bo'lmagan holatda ushlab turadi. Natijada, postabsorbtsiya davrida va ro'za paytida xolesterol sintezi endogen sintez bilan inhibe qilinadi. Agar oziq-ovqat tarkibidagi xolesterin miqdori 2% ga etkazilgan bo'lsa, unda endogen xolesterin sintezi keskin pasaygan. Ammo xolesterin sintezini to'liq to'xtatish sodir bo'lmaydi.
Ovqatdan keladigan xolesterol ta'siri ostida xolesterin biosintezining inhibe darajasi odamdan har kishiga farq qiladi. Bu xolesterolni shakllantirish jarayonlarining individualligini ko'rsatadi. Xolesterol sintezining intensivligini kamaytirish orqali qondagi uning konsentratsiyasini kamaytirish mumkin.
Agar bir tomondan xolesterolni iste'mol qilish va organizmda sintezi, boshqa tomondan safro kislotalari va xolesterolning chiqishi o'rtasidagi muvozanat buzilsa, to'qimalarda va qonda xolesterin kontsentratsiyasi o'zgaradi. Eng jiddiy oqibatlar qonda xolesterin kontsentratsiyasining oshishi (giperkolesterolemiya) bilan bog'liq, ateroskleroz va xolelitiyoz rivojlanishi ehtimolligi oshadi.
Oilaviy giperkolesterolemiya (HCS) - bu shakl ko'proq tarqalgan - har 200 kishiga 1 bemor. HCS-ning meros qilib olingan nuqsoni bu LDL-ni hujayralar tomonidan so'rilishini buzish va shuning uchun LDL katabolizmi tezligining pasayishi. Natijada qonda LDL kontsentratsiyasi, shuningdek xolesterin miqdori oshadi, chunki LDLda bu juda ko'p. Shuning uchun HCS bilan xolesterolni to'qimalarda, xususan terida (ksantomalar), arteriyalar devorlarida cho'ktirish xarakterlidir.
HMG-CoA reduktaza sintezini inhibe qilish
Xolesterol metabolik yo'lining yakuniy mahsuloti. HMG-CoA reduktaz genini transkripsiya tezligini kamaytiradi va shu bilan o'z sintezini inhibe qiladi. Jigar safro kislotalarini xolesteroldan faol ravishda sintez qilmoqda va shuning uchun safro kislotalari HMG-CoA reduktaz genining faolligini inhibe qiladi. HMG-CoA reduktazasi taxminan 3 ta sintezdan so'ng mavjud bo'lganligi sababli, ushbu xolesterin fermenti sintezini inhibe qilish samarali tartibga soluvchi vositadir.
Qidirayotgan narsangizni topmadingizmi? Izlashdan foydalaning:
Xolesterin esterlari almashinuvi
Xolesterol fondida xolesterin va qonda lipoproteinlarda joylashgan xolesterin va xolesterin esterlari mavjud.
II qism Metabolizm va energiya
Xujayralarda xolesterolni esterifikatsiyasi asil-KoA-xolesterol-asiltransferaza (AChAT) ta'sirida sodir bo'ladi:
Acil-CoA + xolesterol - * HS-KoA + Akilxolesterol
Odam hujayralarida asosan linoleyxxolesterin hosil bo'ladi. Erkin xolesteroldan farqli o'laroq, uning hujayra membranalaridagi esterlari juda kam miqdorda bo'ladi va asosan sitozolda lipid tomchilari tarkibida bo'ladi. Esterlarning shakllanishini, bir tomondan, ortiqcha xolesterolni membranalardan tozalash mexanizmi, ikkinchi tomondan, xolesterolni hujayrada saqlash mexanizmi sifatida ko'rib chiqish mumkin. Resurslarni safarbar qilish xolesterin esterlarini gidroliz qiladigan esteraza fermentlari ishtirokida sodir bo'ladi:
Akilxolesterol + H 2O - * Yog 'kislotasi + Xolesterol
Esterlarning sintezi va gidrolizi ko'p hujayralarda uchraydi, lekin ayniqsa buyrak usti korteksining hujayralarida faoldir: bu hujayralarda barcha xolesterolning 80 foizigacha esterlar ishtirok etadi, boshqa hujayralarda esa odatda 20 foizdan kam.
Qonda liponroteinlarda esterlarning shakllanishi lesitin-xolesterolli asiltransferaza (LHAT) ishtirokida ro'y beradi, bu esa atsil qoldig'ining i-pozitsiyasidan xolesterolga o'tishini kataliz qiladi (10.31-rasm) LHAT jigarda hosil bo'ladi, qonga kiradi va lipoproteinlarga birikadi. Turli xil lipoproteinlarni esterifikatsiya qilish darajasi sezilarli darajada farq qiladi va LHATni (asosan apo-AT, shuningdek CI) faollashtiradigan yoki ushbu fermentni (C-II) inhibe qiladigan apolipoproteinlarning mavjudligiga bog'liq. HDLda eng faol LHAT, bunda apo-AT kerak Ölee barcha oqsillarning 2/3. oleyk va linoleik kislotalar Ester hosil yirik coli sharaf. Ester barpo boshqa lipoproteinler HDL nisbatan sekin, bir maromda sodir bo'ladi.
Shakl 10.31. LHAT ta'siri ostida xolesterin esterlarining shakllanishi
LHAT HDL sirt qatlamida lokalizatsiya qilinadi va fosfolipid monolayeridagi xolesterolni substrat sifatida ishlatadi. Bu erda hosil bo'lgan xolesterin esterlari to'liq hidrofobikligi tufayli kam saqlanib qoladi
10-bob. Metabolizm va lipidlarning ishlashi
fosfolipid monolayeri va lipoproteinning lipid yadrosiga botiriladi. Shu bilan birga, xolesterin uchun joy fosfolipidli monolayerda bo'shatiladi, uni xolesterin bilan hujayra membranalari yoki boshqa lipoproteinlar to'ldirishi mumkin. Shunday qilib, HDL LHAT ta'siri natijasida xolesterin tuzog'iga aylanadi.
Safro kislotasi sintezi
Jigarda xolesterolning bir qismi safro kislotalariga aylanadi. Gall kislotalarni xolan kislotasining hosilalari deb hisoblash mumkin (10.32-rasm).
Xolan kislotasi organizmda hosil bo'lmaydi. Gepatotsitlarda xolesterin to'g'ridan-to'g'ri cenodeoxycholic va xolic kislotalarini - birlamchi safro kislotalarini chiqaradi (10.33-rasm, 10.12-rasmga ham qarang).
Xolesterin biosintezi
Xolesterol biosintezi endoplazmatik retikulumda uchraydi. Molekuladagi barcha uglerod atomlarining manbai bu erda yog 'kislotalari sintezidagi kabi sitratning bir qismi sifatida mitoxondriyalardan kelib chiqqan atsetil-SCoA. Xolesterin biosintezi 18 ATP molekulasini va 13 NADPH molekulasini iste'mol qiladi.
Xolesterolning shakllanishi 30 dan ortiq reaktsiyalarda uchraydi, ularni bir necha bosqichda guruhlash mumkin.
1. Mevalonik kislota sintezi.
Dastlabki ikkita sintez reaktsiyasi ketogenez reaktsiyalariga to'g'ri keladi, ammo 3-gidroksi-3-metilglyutaril-ScoA sintezidan keyin ferment fermentga kiradi. gidroksimetil-glutaril-ScoA reduktazasi (HMG-SCOA reduktaz) mevalon kislotasini hosil qiladi.
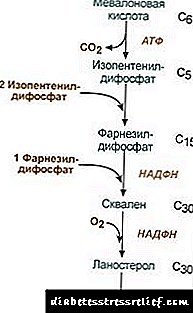 |
Xolesterol sintezining reaktsiyasi sxemasi2. Izopentenil difosfat sintezi. Ushbu bosqichda uchta fosfat qoldig'i mevalon kislotasiga biriktiriladi, keyin u dekarboksillanadi va suvsizlanadi. 3. Izopentenil difosfatning uchta molekulasini birlashtirgandan so'ng, farnesil difosfat sintezlanadi. 4. Ikki farnesil difosfat qoldig'i bog'langanida skalen sintezi sodir bo'ladi. 5. Murakkab reaktsiyalardan so'ng, chiziqli skalol lanosterolga o'tadi. 6. Ortiqcha metil guruhlarini yo'q qilish, molekulani tiklash va izomerizatsiya qilish xolesterinning paydo bo'lishiga olib keladi. Gidroksimetilglyutaril-S-CoA reduktazasining faolligini tartibga solish3. Xolesterin biosintezi tezligi ham kontsentratsiyaga bog'liq o'ziga xos tashuvchi oqsilgidrofobik oraliq sintez metabolitlarini bog'lash va tashishni ta'minlaydi. Siz o'z fikringizni so'rashingiz yoki qoldirishingiz mumkin. Tuzatishning asosiy nuqtasi mevalon kislotasi hosil bo'lish reaktsiyasidir. 1. Allosterik tartibga solish. Xolesterol va jigarda - va safro kislotalari HMG-CoA reduktsinini inhibe qiladi. 2. HMG-CoA reduktaz xolesterin sintezini susaytirish. 3. Faol fosforitlanmagan shaklda HMG-CoA reduktazasining fosforillanish-deposforillanishi bilan tartibga solish. Glyukagon deaktivatsiyaga olib keladi, insulin esa reaktsiyalarning murakkab kaskadida faollashadi. Shunday qilib, changni yutish va postabsorbtsiya holatining o'zgarishi bilan xolesterol sintezi tezligi o'zgaradi. 4. Jigarda HMG-CoA reduktazasini sintez qilish darajasi aniq kunlik tebranishlariga duch keladi: yarim tunda maksimal va ertalab minimal. Xolesterin esterlari almashinuvi Hujayralarda xolesterolni esterifikatsiyasi ta'sir qilganda sodir bo'ladi akil-CoA-xolesterol-asiltransferaza (AHAT): Acil-CoA + Xolesterol ® NS-CoA + Akilxolesterol Hujayralarda asosan linoleyxxolesterin hosil bo'ladi. Esterlar asosan sitozolda lipid tomchilarining bir qismi sifatida joylashgan. Esterlarning shakllanishini, bir tomondan, ortiqcha xolesterolni membranalardan tozalash mexanizmi, ikkinchi tomondan, xolesterolni hujayrada saqlash mexanizmi sifatida ko'rib chiqish mumkin. Zaxiralarni mobilizatsiya qilish fermentlar ishtirokida sodir bo'ladi esterazagidrolizlangan xolesterin esterlari: Akilxolesterol + N2O ® Yog 'kislotasi + xolesterin Esterlarning sintezi va gidrolizlanishi ayniqsa buyrak usti korteksining hujayralarida faollashadi. Qonda lipoproteinlar Ester shakllanishi uning ishtirokida sodir bo'ladi lesitin-xolesterol-asiltransferaza (LHAT), atsil qoldig'ini lesitindan xolesteringa o'tkazilishini katalizlaydi. LHAT jigarda hosil bo'ladi, qonga kiradi va lipoproteinlarga birikadi. HDLda eng faol LHAT, bu erda u sirt qatlamida lokalizatsiya qilinadi. Bu erda hosil bo'lgan xolesterin esterlari hidrofobik bo'lib, lipid yadrosiga botiriladi. Fosfolipid monolayerida xolesterin uchun bo'sh joy mavjud bo'lib, u xolesterinni hujayra membranalari yoki boshqa lipoproteinlardan to'ldirishi mumkin. Shunday qilib, HDL LHAT ta'siri natijasida xolesterin tuzog'iga aylanadi. Safro kislotasi sintezi Jigarda xolesterolning bir qismi safro kislotalariga aylanadi. Safro kislotalarini xolan kislotasining hosilalari sifatida ko'rib chiqish mumkin. Xolan kislotasi organizmda hosil bo'lmaydi. Xolesteroldan gepatotsitlarda birlamchi safro kislotalari hosil bo'ladi - cenodeoxycholik va xolerik. Ichak florasi fermenti ta'sirida ichakka safro chiqarilgandan so'ng ulardan ikkilamchi safro kislotalari hosil bo'ladi - litologik va deoksikolik. Ular ichaklardan so'riladi, portal venadagi qon jigarga, keyin safroga kiradi. Safro asosan konjugatsiyalangan safro kislotalarini, ya'ni ularning glitsin yoki taurin bilan birikmalarini o'z ichiga oladi. Safro safro kislotalarining kontsentratsiyasi taxminan 1% ni tashkil qiladi. Safro kislotalarining asosiy qismi ishtirok etadi gepatoenterik qon aylanishi.Safro kislotalarining ozgina qismi (kuniga taxminan 0,5 g) najas bilan chiqariladi. Bu jigarda yangi safro kislotalarining sintezi bilan qoplanadi, safro kislotasi fondi taxminan 10 kun ichida yangilanadi. Xolesterol asosan ichak orqali chiqariladi. U ichakka oziq-ovqat va safro qismi sifatida kiradi. Qonda so'rilgan xolesterol safrodan kelib chiqadigan fraktsiyani o'z ichiga oladi (endogen xolesterinjigarda sintez qilinadi) va oziq-ovqatdan olingan fraktsiya (ekzogen xolesterin) Xolesterolni to'qimalardan olib tashlash uning jigarda safro kislotalariga oksidlanishi, so'ngra najas bilan chiqishi (kuniga 0,5 g) va o'zgarishsiz xolesterol (najas bilan) chiqarilishi bilan sodir bo'ladi. Statsionar holatda: (Xolesterinoxiri + Xolesterinsobiq) - (Xolesterineksklyuziv + Safro kislotalarieksklyuziv) = 0 Agar bu muvozanat buzilsa, to'qimalarda va qonda xolesterin kontsentratsiyasi o'zgaradi. Qonda xolesterolni ko'payishi - giperkolesterolemiya. Bu ateroskleroz va safro tosh kasalligi ehtimolini oshiradi. LIPID BIRJANING TARTIBI Lipit metabolizmi markaziy asab tizimi tomonidan boshqariladi. Uzoq muddatli salbiy hissiy stress, katekolaminlarni qonga chiqarilishini ko'payishi sezilarli vazn yo'qotishiga olib kelishi mumkin. Harakat glyukagon lipolitik tizimda katekolaminlarning ta'siriga o'xshash. Adrenalin va norepinefrin to'qima lipazasi faolligini va yog 'to'qimasida lipoliz tezligini oshiradi, natijada qon plazmasidagi yog' kislotalari miqdori ortadi. Insulin adrenalin va glyukagonning lipolizga va yog 'kislotalarini safarbar qilishga teskari ta'sir ko'rsatadi. O'sish gormoni asenilat siklaza sintezini keltirib chiqaradigan lipolizni rag'batlantiradi. Gipofiz gipofunktsiyasi tanadagi yog 'to'planishiga olib keladi (gipofiz semirib ketishi). Tiroksin, jinsiy gormonlarlipid metabolizmiga ham ta'sir qiladi. Hayvonlarda jinsiy bezlarni olib tashlash ortiqcha yog'larning to'planishiga olib keladi. LIPID METABOLIK DISORDERLARI Xolesterol hayvonlar organizmiga xos bo'lgan steroiddir. Inson tanasida uning shakllanishidagi asosiy joy jigardir, unda 50% xolesterin sintez qilinadi, 15-20% ingichka ichakda hosil bo'ladi, qolgan qismi terida, adrenal korteksda va gonadlarda sintezlanadi. Xolesterin fondi shakllanish manbalari va uni sarflash usullari 22.1-rasmda keltirilgan. Shakl 22.1. Tanadagi xolesterolning shakllanishi va tarqalishi. Odam tanasining xolesterolini (umumiy miqdori taxminan 140 g) shartli ravishda uchta basseynga bo'lish mumkin: 30 g) tez almashinish, ichak devoridagi xolesterol, qon plazmasi, jigar va boshqa parenximal organlardan iborat, yangilanish 30 kun ichida (kuniga 1 g), 50 g), asta-sekin boshqa organlar va to'qimalarning xolesterolini almashtirish, 60 g), orqa miya va miya, biriktiruvchi to'qima juda sekin almashinadigan xolesterin, yangilanish darajasi yillar davomida hisoblab chiqilgan. Xolesterol sintezi hujayralar sitosolida sodir bo'ladi. Bu inson organizmidagi metabolik yo'llarning eng uzun yo'llaridan biridir. U 3 bosqichda davom etadi: birinchisi mevalonik kislota, ikkinchisi skalen (30 uglerod atomidan iborat chiziqli uglevodorod tuzilishi) hosil bo'lishi bilan tugaydi. Uchinchi bosqichda skalen lanosterol molekulasiga aylanadi, keyin lanosterolni xolesteringa aylantiradigan 20 ta ketma-ket reaktsiyalar mavjud. Ba'zi to'qimalarda xolesterin gidroksil guruhi esterlarni hosil qiladi. Reaktsiya hujayra ichidagi AHAT fermenti tomonidan katalizlanadi (acylCoA: xolesterol asiltransferaza). Esterifikatsiya reaktsiyasi LHAT fermenti (lesitin: xolesterin asiltransferaza) joylashgan HDLda ham uchraydi. Xolesterin esterlari - bu qon orqali tashiladigan yoki hujayralarga joylashadigan shakl. Qonda xolesterolning taxminan 75% esterlar shaklida bo'ladi. Xolesterol sintezi jarayonning asosiy fermenti - 3-gidroksi-3-metilglyutaril-CoA reduktazasi (HMG-CoA reduktazasi) ning faolligi va miqdoriga ta'sir qilish orqali tartibga solinadi. Bunga ikki yo'l bilan erishiladi: 1. HMG-CoA reduktazasining fosforillanishi / deposforillanishi. Insulin HMG-CoA reduktazasining deposforlanishini rag'batlantiradi va shu bilan uni faol holatga o'tkazadi. Shuning uchun, assimilyatsiya davrida xolesterol sintezi kuchayadi. Ushbu davrda asetil-CoA sintezi uchun boshlang'ich substratning mavjudligi ham ortadi. Glyukagon teskari ta'sirga ega: oqsil kinazasi orqali HMG-CoA reduktazasining fosforlanishini rag'batlantiradi va uni faol bo'lmagan holatga keltiradi. Natijada, postabsorbtsiya davrida va ro'za paytida xolesterin sintezi inhibe qilinadi. 2. HMG-CoA reduktaza sintezini inhibe qilish.Xolesterol (metabolik yo'lning yakuniy mahsuloti) HMG-CoA reduktaz genining transkripsiya tezligini pasaytiradi va shu bilan o'z sintezini inhibe qiladi, safro kislotalari ham shunga o'xshash ta'sirga olib keladi. Qonda xolesterolni tashish preparatning bir qismi sifatida amalga oshiriladi. LP to'qimalarda ekzogen xolesterolni ta'minlaydi, uning organlar orasidagi oqishini va tanadan chiqarilishini aniqlaydi. Ekzogen xolesterin jigarga qoldiq ChM tarkibida etkazib beriladi. U erda sintez qilingan endogen xolesterin bilan birgalikda umumiy fond hosil bo'ladi. Gepatotsitlarda TAG va xolesterin VLDL ichiga joylashadi va ushbu shaklda qonga kiradi. Qonda VLDL LG-lipaz ta'siri ostida, TAG ni glitserin va yog 'kislotalariga gidroliz qiladi, avval LSPPga, so'ngra 55% gacha xolesterol va uning esterlariga ega bo'lgan LDLga aylantiriladi. LDL - bu xolesterolning asosiy transport shakli bo'lib, u to'qimalarga etkaziladi (xolesterinning 70 foizi va qon tarkibidagi esterlar LDL tarkibiga kiradi). Qondan LDL jigarga (75% gacha) va ularning yuzasida LDL retseptorlari bo'lgan boshqa to'qimalarga kiradi. Agar hujayraga kiradigan xolesterin miqdori uning ehtiyojidan oshsa, u holda LDL retseptorlari sintezi bostiriladi, bu qondan xolesterin oqimini kamaytiradi. Hujayrada bo'sh xolesterin kontsentratsiyasining pasayishi bilan, aksincha, retseptorlarning sintezi faollashadi. LDL retseptorlari sintezini tartibga solishda gormonlar ishtirok etadi: insulin, triiodothyronine va jinsiy gormonlar retseptorlarning shakllanishini oshiradi va glyukokortikoidlar kamayadi. "Xolesterolni qaytarish transporti" deb ataladigan joyda, ya'ni. xolesterolni jigarga qaytarilishini ta'minlaydigan yo'l, HDL asosiy rol o'ynaydi. Ular jigarda deyarli xolesterin va TAGni o'z ichiga olmaydigan etuk bo'lmagan prekursorlar shaklida sintezlanadi. Qondagi HDL prekursorlari xolesterolga to'yingan bo'lib, uni boshqa LP va hujayra membranalaridan oladi. Xolesterolni HDL ga o'tkazish ularning yuzasida joylashgan LHAT fermentini o'z ichiga oladi. Ushbu ferment yog 'kislotasi qoldig'ini fosfatidilkolin (lesitin) dan xolesteringa qo'shadi. Natijada HDL ichida harakatlanadigan xolesterol esterining hidrofobik molekulasi hosil bo'ladi. Shunday qilib, xolesterol bilan boyitilmagan HDL HDL 3 ga aylanadi - etuk va kattaroq zarralar. HDL 3 VLDL va STD tarkibidagi TAG uchun xolesterin esterlarini lipoproteinlar orasidagi xolesterin esterlarini o'tkazadigan ma'lum bir protein ishtirokida almashtiring. Bunday holda, HDL 3 HDL2 ga aylanadilar, ularning kattaligi TAG to'planishi natijasida oshadi. LLD-lipaz ta'sirida VLDL va STDL asosan jigarga xolesterolni etkazib beradigan LDLga aylantiriladi. Xolesterolning oz qismi HDL2 va HDL jigariga etkaziladi. Safro kislotalari sintezi. Jigarda kuniga 500-700 mg safro kislotalari xolesteroldan sintez qilinadi. Ularning shakllanishiga gidroksilazalar ishtirokidagi gidroksil guruhlari va xolesterin yon zanjirining qisman oksidlanish reaktsiyalari kiradi (22.2-rasm): Shakl 22.2. Safro kislotasining shakllanishi. Birinchi sintez reaktsiyasi - 7-a-gidroksikolesterolning shakllanishi - bu tartibga soluvchi. Ushbu reaktsiyani katalizlaydigan fermentning faoliyati yo'lning oxirgi mahsuloti, safro kislotalari tomonidan inhibe qilinadi. Yana bir tartibga solish mexanizmi bu fermentning fosforillanishi / deposforillanishi (7-a-gidroksilazaning fosforlangan shakli faol). Ferment miqdorini o'zgartirish orqali ham tartibga solish mumkin: xolesterin 7-a-gidroksilaza genining transkripsiyasini keltirib chiqaradi va safro kislotalari bosim o'tkazadilar. Tiroid gormonlari 7-a-gidroksilaza sintezini qo'zg'atadi va estrogen bosimini pasaytiradi. Estrogenning safro kislotalari sinteziga ta'siri, o't pufagi kasalligi nega ayollarda erkaklarga qaraganda 3-4 baravar tez-tez uchrab turishini tushuntiradi. Xolesteroldan hosil bo'lgan kolik va cenodeoxycholik kislotalar "birlamchi safro kislotalari" deb ataladi. Ushbu kislotalarning asosiy qismi konjugatsiyaga uchraydi - safro kislotasining karboksil guruhiga glisin yoki taurin molekulalari qo'shilishi. Konjugatsiya safro kislotalarining faol shakli - CoA hosilalari hosil bo'lishidan boshlanadi, so'ngra taurin yoki glisin birikadi va natijada 4 ta konjugatlar paydo bo'ladi: taurokolik va taurohenodeoxycholik, glikokolik va glikoxenodeoksikolik kislotalar. Ular asl safro kislotalariga qaraganda ancha kuchli emulsifikatorlardir. Glisin bilan kon'yugatlar tauringa qaraganda 3 baravar ko'proq hosil bo'ladi, chunki tanadagi taurin miqdori cheklangan. Ichakda bakterial fermentlar ta'sirida oz miqdordagi birlamchi safro kislotalari konjugatlari ikkilamchi safro kislotalariga aylanadi. Xoldan hosil bo'lgan deoksikolik kislota va deoksikolikdan hosil bo'lgan litoxolik kislota kamroq eriydi va ichaklarda sekinroq so'riladi. Ichakka kiradigan safro kislotalarining qariyb 95% jigarga portal tomir orqali qaytadi, so'ngra yana safroga chiqariladi va yog'larning emulsifikatsiyasida qayta ishlatiladi. Safro kislotalarining bu yo'li enteroepatik aylanish deb ataladi. Najas bilan, ikkilamchi safro kislotalari asosan chiqariladi. O't toshlari kasalligi (xolelitiyoz) bu patologik jarayon bo'lib, uning tarkibida xolesterol bo'lgan o't pufagida toshlar paydo bo'ladi. Xolesterolni safroga chiqishi safro kislotalari va gidrofob xolesterin molekulalarini mikroelement holatida ushlab turuvchi fosfolipidlarning mutanosib ravishda chiqarilishi bilan birga bo'lishi kerak. Safro tarkibidagi safro kislotalari va xolesterin miqdorining o'zgarishiga olib keladigan sabablar: xolesterolga boy oziq-ovqat, yuqori kaloriyali ovqatlanish, o't pufagida safro turg'unligi, enteroepatik qon aylanishining buzilishi, safro kislotalari sintezining buzilishi, o't pufagi infektsiyasi. Xolelitiyoz kasalligi bo'lgan bemorlarning ko'pchiligida xolesterin sintezi kuchayadi va undan safro kislotalari sintezi sekinlashadi, bu esa safro ichiga tushgan xolesterin va safro kislotalari sonining nomutanosibligiga olib keladi. Natijada, o't pufagida xolesterin asta-sekin qattiqlashadigan yopishqoq cho'kma hosil bo'ladi. Ba'zida u bilirubin, oqsillar va kaltsiy tuzlari bilan singdiriladi. Toshlar faqat xolesterol (xolesterin toshlari) yoki xolesterin, bilirubin, oqsillar va kaltsiy aralashmasidan iborat bo'lishi mumkin. Xolesterol toshlari odatda oq rangga ega, aralash toshlar turli xil soyalarda jigarrang. Tosh paydo bo'lishining dastlabki bosqichida chenodeoxycholik kislota dori sifatida ishlatilishi mumkin. O't pufagida xolesterol toshlarini asta-sekin eritib yuboradi, ammo bu bir necha oy davom etadigan sekin jarayon. Ateroskleroz - bu qon tomir devorining ichki yuzasida aterogen plaklarning paydo bo'lishi bilan tavsiflangan patologiya. Bunday patologiyaning rivojlanishining asosiy sabablaridan biri bu xolesterolni ovqatdan olish, uning sintezi va organizmdan chiqishi o'rtasidagi muvozanatni buzishdir. Aterosklerozli bemorlarda LDL va VLDL kontsentratsiyalari ko'paygan. HDL kontsentratsiyasi va aterosklerozning rivojlanish ehtimoli o'rtasida teskari aloqa mavjud. Bu LDLning to'qimalarda xolesterolni va to'qimalardan HDLni tashuvchisi sifatida ishlash kontseptsiyasiga mos keladi. Ateroskleroz rivojlanishining asosiy metabolik "sharti" bu giperkolesterolemiya. (qondagi yuqori xolesterin). Giperkolesterolemiya rivojlanadi: 1. xolesterol, uglevodlar va yog'larning ortiqcha iste'mol qilinishi natijasida 2. LDL yoki apoB-100 retseptorlari tuzilishidagi irsiy nuqsonlardan, shuningdek apoB-100 sintezi yoki sekretsiyasining oshishidan iborat bo'lgan genetik moyillik (oilaviy estrodiol giperlipidemiya holatida, unda qonda xolesterin va TAG ko'tarilgan). Ateroskleroz rivojlanish mexanizmlarida preparatning modifikatsiyasi muhim rol o'ynaydi. LDLdagi lipidlar va oqsillarning normal tuzilishidagi o'zgarishlar ularni tanaga begona qiladi va shuning uchun fagotsitlar tomonidan ushlanishi uchun qulayroqdir. Giyohvand modifikatsiyasi bir necha mexanizmlar yordamida yuzaga kelishi mumkin: 1. Qondagi glyukoza kontsentratsiyasi oshganda hosil bo'lgan oqsillarning glikozillanishi 2. peroksid modifikatsiyasi lipoproteinlardagi lipidlarning o'zgarishiga va apoB-100 tarkibiga kirishi, 3. LP-antikorining otoimmun komplekslarining shakllanishi (o'zgartirilgan dorilar otoantikorlarning paydo bo'lishiga olib kelishi mumkin). Modifikatsiyalangan LDL makrofaglar tomonidan so'riladi. Bu xolesterin so'rilgan xolesterin miqdori bilan tartibga solinmaydi, chunki u maxsus retseptorlar orqali hujayralarga kirganda, shuning uchun makrofaglar xolesterin bilan haddan tashqari ko'payib, subendotelial bo'shliqqa kirib boradigan "ko'pikli hujayralar" ga aylanadi. Bu qon tomirlari devorida lipid dog'lari yoki chiziqlar paydo bo'lishiga olib keladi. Ushbu bosqichda qon tomir endoteliy o'z tuzilishini saqlab turishi mumkin. Ko'pik hujayralari sonining ko'payishi bilan endotelial shikastlanish paydo bo'ladi. Zarar trombotsitlarning faollashishiga yordam beradi. Natijada, ular trombotsitlar to'planishini rag'batlantiruvchi tromboksanni ajratadilar va silliq mushak hujayralari ko'payishini rag'batlantiradigan trombotsitlardan kelib chiqqan o'sish omilini ishlab chiqarishni boshlaydilar. Ikkinchisi medialdan arteriya devorining ichki qavatiga o'tadi va shu bilan blyashka o'sishiga yordam beradi. Keyinchalik blyashka tolali to'qima bilan o'sib chiqadi, tolali membrananing ostidagi hujayralar nekrotikdir va xolesterin hujayralararo bo'shliqqa joylashadi. Rivojlanishning so'nggi bosqichlarida blyashka kaltsiy tuzlari bilan singdiriladi va juda zich bo'ladi. Blyashka hududida ko'pincha qon pıhtıları shakllanadi, bu tomirning lümenini to'sib qo'yadi, bu esa tegishli to'qima joyida o'tkir qon aylanishining buzilishiga va yurak xurujining rivojlanishiga olib keladi. Xolesterin sintezini tartibga solish - uning asosiy fermenti (HMG-CoA reduktaza) turli yo'llar bilan amalga oshiriladi. HMG reduktazasining fosforillanishi / deposforillanishi. Insulin / glyukagon nisbati oshishi bilan bu ferment fosforilatlanadi va faol holatga o'tadi. Insulinning ta'siri 2 ta ferment orqali amalga oshiriladi. HMG-CoA redüktaz kinaz fosfataza, bu kinazani faol bo'lmagan defosforillangan holatga aylantiradi: Fosfotaza HMG-CoA reduktazasini uni dehosforlangan faol holatga o'tkazish orqali. Ushbu reaktsiyalarning natijasi HMG-CoA reduktazasining dehosforlangan faol shaklini shakllantirishdir. Shunday qilib, assimilyatsiya davrida xolesterin miqdori oshadi. Ushbu davrda xolesterin - asetil - CoA sintezi uchun dastlabki substratning mavjudligi oshadi (uglevodlar va yog'larni o'z ichiga olgan oziq-ovqatlarni iste'mol qilish natijasida, chunki CoA asetil glyukoza va yog 'kislotalari parchalanishi jarayonida hosil bo'ladi). Postabsorbent holatda, proteingenaza A orqali glyukagon HMG - CoA - reduktsazning fosforlanishini rag'batlantiradi va uni faol bo'lmagan holatga aylantiradi. Ushbu harakat glyukagon bir vaqtning o'zida HMG-CoA reduktazasining fosforlanishini va inaktivatsiyasini rag'batlantiradi va shu bilan HMG-CoA redüktazasini fosforillangan faol bo'lmagan holatda ushlab turadi. Natijada, postabsorbtsiya davrida va ro'za paytida xolesterol sintezi endogen sintez bilan inhibe qilinadi. Agar oziq-ovqat tarkibidagi xolesterin miqdori 2% ga etkazilgan bo'lsa, unda endogen xolesterin sintezi keskin pasaygan. Ammo xolesterin sintezini to'liq to'xtatish sodir bo'lmaydi. Ovqatdan keladigan xolesterol ta'siri ostida xolesterin biosintezining inhibe darajasi odamdan har kishiga farq qiladi. Bu xolesterolni shakllantirish jarayonlarining individualligini ko'rsatadi. Xolesterol sintezining intensivligini kamaytirish orqali qondagi uning konsentratsiyasini kamaytirish mumkin. Agar bir tomondan xolesterolni iste'mol qilish va organizmda sintezi, boshqa tomondan safro kislotalari va xolesterolning chiqishi o'rtasidagi muvozanat buzilsa, to'qimalarda va qonda xolesterin kontsentratsiyasi o'zgaradi. Eng jiddiy oqibatlar qonda xolesterin kontsentratsiyasining oshishi (giperkolesterolemiya) bilan bog'liq, ateroskleroz va xolelitiyoz rivojlanishi ehtimolligi oshadi. Oilaviy giperkolesterolemiya (HCS) - bu shakl ko'proq tarqalgan - har 200 kishiga 1 bemor. HCS-ning meros qilib olingan nuqsoni bu LDL-ni hujayralar tomonidan so'rilishini buzish va shuning uchun LDL katabolizmi tezligining pasayishi. Natijada qonda LDL kontsentratsiyasi, shuningdek xolesterin miqdori oshadi, chunki LDLda bu juda ko'p. Shuning uchun HCS bilan xolesterolni to'qimalarda, xususan terida (ksantomalar), arteriyalar devorlarida cho'ktirish xarakterlidir. HMG-CoA reduktaza sintezini inhibe qilish Xolesterol metabolik yo'lining yakuniy mahsuloti. HMG-CoA reduktaz genini transkripsiya tezligini kamaytiradi va shu bilan o'z sintezini inhibe qiladi. Jigar safro kislotalarini xolesteroldan faol ravishda sintez qilmoqda va shuning uchun safro kislotalari HMG-CoA reduktaz genining faolligini inhibe qiladi. HMG-CoA reduktazasi taxminan 3 ta sintezdan so'ng mavjud bo'lganligi sababli, ushbu xolesterin fermenti sintezini inhibe qilish samarali tartibga soluvchi vositadir.
|